Citation: Tara L. Renbarger, Jennifer M. Baker, W. Matthew Sattley. Slow and steady wins the race: an examination of bacterial persistence[J]. AIMS Microbiology, 2017, 3(2): 171-185. doi: 10.3934/microbiol.2017.2.171

| [1] | Sengupta S, Chattopadhyay MK, Grossart HP (2013) The multifaceted roles of antibiotics and antibiotic resistance in nature. Front Microbiol 4: 47. |
| [2] | Centers for Disease Control and Prevention (2013) Antibiotic resistance threats in the United States, 2013. |
| [3] | Abraham EP, Chain E (1940) An enzyme from bacteria able to destroy penicillin. Nature 146: 837. |
| [4] | Bigger JW (1944) Treatment of staphylococcal infections with penicillin by intermittent sterilisation. Lancet 2: 497–500. |
| [5] |
Moyed HS, Broderick SH (1986) Molecular cloning and expression of hipA, a gene of Escherichia coli K-12 that affects frequency of persistence after inhibition of murein synthesis. J Bacteriol 166: 399–403. doi: 10.1128/jb.166.2.399-403.1986

|
| [6] | Centers for Disease Control and Prevention (2016) Newly reported gene, mcr-1, threatens last-resort antibiotics. |
| [7] |
Mwangi MM, Wu SW, Zhou Y, et al. (2007) Tracking the in vivo evolution of multidrug resistance in Staphylococcus aureus by whole-genome sequencing. Proc Natl Acad Sci USA 104: 9451–9456. doi: 10.1073/pnas.0609839104
|
| [8] | Liu B, Pop M (2009) ARDB-Antibiotic Resistance Genes Database. Nucleic Acids Res 37(Database issue): D443–D447. |
| [9] |
Kussell E, Kishony R, Balaban NQ, et al. (2005) Bacterial persistence: a model of survival in changing environments. Genetics 169: 1807–1814. doi: 10.1534/genetics.104.035352
|
| [10] | de Jong IG, Haccou P, Kuipers OP (2011) Bet hedging or not? A guide to proper classification of microbial survival strategies. Bioessays 33: 215–223. |
| [11] | Moyed HS, Bertrand KP (1983) hipA, a newly recognized gene of Escherichia coli K-12 that affects frequency of persistence after inhibition of murein synthesis.J Bacteriol 155: 768–775. |
| [12] |
Korch SB, Henderson TA, Hill TM (2003) Characterization of the hipA7 allele of Escherichia coli and evidence that high persistence is governed by (p)ppGpp synthesis.Mol Microbiol 50: 1199–1213. doi: 10.1046/j.1365-2958.2003.03779.x
|
| [13] |
Spoering AL, Lewis K (2001) Biofilms and planktonic cells of Pseudomonas aeruginosa have similar resistance to killing by antimicrobials.J Bacteriol 183: 6746–6751. doi: 10.1128/JB.183.23.6746-6751.2001
|
| [14] |
Keren I, Kaldalu N, Spoering A, et al. (2004) Persister cells and tolerance to antimicrobials. FEMS Microbiol Lett 230: 13–18. doi: 10.1016/S0378-1097(03)00856-5
|
| [15] |
Mulcahy LR, Burns JL, Lory S, et al. (2010) Emergence of Pseudomonas aeruginosa strains producing high levels of persister cells in patients with cystic fibrosis. J Bacteriol 192: 6191–6199. doi: 10.1128/JB.01651-09
|
| [16] |
Spoering AL, Vulic M, Lewis K (2006) GlpD and PlsB participate in persister cell formation in Escherichia coli. J Bacteriol 188: 5136–5144. doi: 10.1128/JB.00369-06
|
| [17] | Hofsteenge N, van Nimwegen E, Silander OK (2013) Quantitative analysis of persister fractions suggests different mechanisms of formation among environmental isolates of E. coli. BMC Microbiol 13: 25. |
| [18] |
Orman MA, Brynildsen MP (2013) Dormancy is not necessary or sufficient for bacterial persistence. Antimicrob Agents Chemother 57: 3230–3239. doi: 10.1128/AAC.00243-13
|
| [19] |
Brouqui P, Bacellar F, Baranton G, et al. (2004) Guidelines for the diagnosis of tick-borne bacterial diseases in Europe. Clin Microbiol Infect 10: 1108–1132. doi: 10.1111/j.1469-0691.2004.01019.x
|
| [20] | Lewis K (2008) Multidrug tolerance of biofilms and persister cells.Curr Top Microbiol Immunol 322: 107–131. |
| [21] |
Brooun A, Liu S, Lewis K (2000) A dose-response study of antibiotic resistance in Pseudomonas aeruginosa biofilms.Antimicrob Agents Chemother 44: 640–646. doi: 10.1128/AAC.44.3.640-646.2000
|
| [22] |
Lewis K (2001) Riddle of biofilm resistance.Antimicrob Agents Chemother 45: 999–1007. doi: 10.1128/AAC.45.4.999-1007.2001
|
| [23] |
Luidalepp H, Jõers A, Kaldalu N, et al. (2011) Age of inoculum strongly influences persister frequency and can mask effects of mutations implicated in altered persistence.J Bacteriol 193: 3598–3605. doi: 10.1128/JB.00085-11
|
| [24] |
Balaban NQ, Merrin J, Chait R, et al. (2004) Bacterial persistence as a phenotypic switch.Science 305: 1622–1625. doi: 10.1126/science.1099390
|
| [25] |
Jõers A, Kaldalu N, Tenson T (2010) The frequency of persisters in Escherichia coli reflects the kinetics of awakening from dormancy.J Bacteriol 192: 3379–3384. doi: 10.1128/JB.00056-10
|
| [26] |
Roostalu J, Jõers A, Luidalepp H, et al. (2008) Cell division in Escherichia coli cultures monitored at single cell resolution.BMC Microbiol 8: 68. doi: 10.1186/1471-2180-8-68
|
| [27] | Field MF, Lichstein HC (1958) Growth stimulating effect of autoclaved glucose media and its relationship to the CO2 requirement of propionibacteria.J Bacteriol 76: 485–490. |
| [28] | Rohmer L, Didier H, Miller SI (2011) Are pathogenic bacteria just looking for food? Metabolism and microbial pathogenesis. Trends Microbiol 19: 341–348. |
| [29] |
Amato S, Brynildsen M (2014) Nutrient transitions are a source of persisters in Escherichia coli biofilms. PLoS One 9: e93110. doi: 10.1371/journal.pone.0093110
|
| [30] |
Kotte O, Volkmer B, Radzikowski JL, et al. (2014) Phenotypic bistability in Escherichia coli's central carbon metabolism. Mol Syst Biol 10: 736. doi: 10.15252/msb.20135022
|
| [31] |
Radzikowski JL, Vedelaar S, Siegel D, et al. (2016) Bacterial persistence in an active σS stress response to metabolic flux limitation. Mol Syst Biol 12: 882. doi: 10.15252/msb.20166998
|
| [32] |
Gentry DR, Hernandez VJ, Nguyen LH, et al. (1993) Synthesis of the stationary-phase sigma factor sigma s is positively regulated by ppGpp. J Bacteriol 175: 7982–7989. doi: 10.1128/jb.175.24.7982-7989.1993
|
| [33] |
Baillie GS, Douglas LJ (1999) Candida biofilms and their susceptibility to antifungal agents.Methods Enzymol 310: 644–656. doi: 10.1016/S0076-6879(99)10050-8
|
| [34] |
Costerton JW, Stewart PS, Greenberg EP (1999) Bacterial biofilms: a common cause of persistent infections.Science 284: 1318–1322. doi: 10.1126/science.284.5418.1318
|
| [35] | Stewart PS (2015) Antimicrobial Tolerance in Biofilms.Microbiol Spectr 3. |
| [36] | Dalebroux ZD, Swanson MS (2012) ppGpp: magic beyond RNA polymerase.Nat Rev Microbiol 10: 203–212. |
| [37] |
Maisonneuve E, Castro-Camargo M, Gerdes K (2013) (p)ppGpp controls bacterial persistence by stochastic induction of toxin-antitoxin activity.Cell 154: 1140–1150. doi: 10.1016/j.cell.2013.07.048
|
| [38] |
Wood TK, Knabel SJ, Kwan BW (2013) Bacterial persister cell formation and dormancy.Appl Environ Microbiol 79: 7116–7121. doi: 10.1128/AEM.02636-13
|
| [39] |
Jayaraman R (2008) Bacterial persistence: some new insights into an old phenomenon.J Biosci 33: 795–805. doi: 10.1007/s12038-008-0099-3
|
| [40] |
Black DS, Kelly AJ, Mardis MJ, et al. (1991) Structure and organization of hip, an operon that affects lethality due to inhibition of peptidoglycan or DNA synthesis.J Bacteriol 173: 5732–5739. doi: 10.1128/jb.173.18.5732-5739.1991
|
| [41] |
Black DS, Irwin B, Moyed HS (1994) Autoregulation of hip, an operon that affects lethality due to inhibition of peptidoglycan or DNA synthesis.J Bacteriol 176: 4081–4091. doi: 10.1128/jb.176.13.4081-4091.1994
|
| [42] |
Schumacher MA, Piro KM, Xu W, et al. (2009) Molecular mechanisms of HipA-mediated multidrug tolerance and its neutralization by HipB.Science 323: 396–401. doi: 10.1126/science.1163806
|
| [43] |
Hansen S, Vulić M, Min J, et al. (2012) Regulation of the Escherichia coli HipBA toxin-antitoxin system by proteolysis. PLoS One 7: e39185. doi: 10.1371/journal.pone.0039185
|
| [44] |
Germain E, Castro-Roa D, Zenkin N, et al. (2013) Molecular mechanism of bacterial persistence by HipA. Molecular Cell 52: 248–254. doi: 10.1016/j.molcel.2013.08.045
|
| [45] | English BP, Hauryliuk V, Sanamrad A, et al. (2011) Single-molecule investigation of the stringent response machinery in living bacterial cells. Proc Natl Acad Sci USA 108: e365–373. |
| [46] |
Keren I, Shah D, Spoering A, et al. (2004) Specialized persister cells and the mechanism of multidrug tolerance in Escherichia coli.J Bacteriol 186: 8172–8180. doi: 10.1128/JB.186.24.8172-8180.2004
|
| [47] |
Rotem E, Loinger A, Ronin I, et al. (2010) Regulation of phenotypic variability by a threshold-based mechanism underlies bacterial persistence.Proc Natl Acad Sci USA 107: 12541–12546. doi: 10.1073/pnas.1004333107
|
| [48] |
Korch SB, Hill TM (2006) Ectopic overexpression of wild-type and mutant hipA genes in Escherichia coli: effects on macromolecular synthesis and persister formation.J Bacteriol 188: 3826–3836. doi: 10.1128/JB.01740-05
|
| [49] |
Christensen SK, Gerdes K (2003) RelE toxins from Bacteria and Archaea cleave mRNAs on translating ribosomes, which are rescued by tmRNA.Mol Microbiol 48: 1389–1400. doi: 10.1046/j.1365-2958.2003.03512.x
|
| [50] |
Spoering AL, Vulic M, Lewis K (2006) GlpD and PlsB participate in persister cell formation in Escherichia coli.J Bacteriol 188: 5136–5144. doi: 10.1128/JB.00369-06
|
| [51] |
Pedersen K, Zavialov AV, Pavlov MY, et al. (2003) The bacterial toxin RelE displays codon-specific cleavage of mRNAs in the ribosomal A site.Cell 112: 131–140. doi: 10.1016/S0092-8674(02)01248-5
|
| [52] |
Zhang Y, Zhang H, Hoeflich KP, et al. (2003) MazF cleaves cellular mRNAs specifically at ACA to block protein synthesis in Escherichia coli.Mol Cell 12: 913–923. doi: 10.1016/S1097-2765(03)00402-7
|
| [53] |
Shah D, Zhang Z, Khodursky AB, et al. (2006) Persisters: a distinct physiological state of E. coli.BMC Microbiol 6: 53. doi: 10.1186/1471-2180-6-53
|
| [54] |
Pandey DP, Gerdes K (2005) Toxin-antitoxin loci are highly abundant in free-living but lost from host-associated prokaryotes. Nucleic Acids Res 33: 966–976. doi: 10.1093/nar/gki201
|
| [55] |
Hu Y, Kwan BW, Osbourne DO, et al. (2015) Toxin YafQ increases persister cell formation by reducing indole signaling.Environ Microbiol 17: 1275–1285. doi: 10.1111/1462-2920.12567
|
| [56] |
Wang D, Ding X, Rather PN (2001) Indole can act as an extracellular signal in Escherichia coli. J Bacteriol 183: 4210–4216. doi: 10.1128/JB.183.14.4210-4216.2001
|
| [57] |
Aizenman E, Engelberg-Kulka H, Glaser G (1996) An Escherichia coli chromosomal "addiction module" regulated by guanosine [corrected] 3',5'-bispyrophosphate: a model for programmed bacterial cell death.Proc Natl Acad Sci USA 93: 6059–6063. doi: 10.1073/pnas.93.12.6059
|
| [58] |
Reed JL, Vo TD, Schilling CH, et al. (2003) An expanded genome-scale model of Escherichia coli K-12 (iJR904 GSM/GPR).Genome Biol 4: R54. doi: 10.1186/gb-2003-4-9-r54
|
| [59] |
Ren D, Bedzyk LA, Thomas SM, et al. (2004) Gene expression in Escherichia coli biofilms.Appl Microbiol Biotechnol 64: 515–524. doi: 10.1007/s00253-003-1517-y
|
| [60] | Gonzalez Barrios AF, Zuo R, Hashimoto Y, et al. (2006) Autoinducer 2 controls biofilm formation in Escherichia coli through a novel motility quorum-sensing regulator (MqsR, B3022).J Bacteriol 188: 305–316. |
| [61] |
Kasari V, Krug K, Margus T, et al. (2010) The Escherichia coli mqsR and ygiI genes encode a new toxin-antitoxin pair. J Bacteriol 192: 2908–2919. doi: 10.1128/JB.01266-09
|
| [62] | Li Y, Zhang Y (2007) PhoU is a persistence switch involved in persister formation and tolerance to multiple antibiotics and stresses in Escherichia coli.Antimicrob Agents Chemother 51: 2092–2099. |
| [63] |
Ma C, SIM S, Shi W, et al. (2010) Energy production genes sucB and ubiF are involved in persister survival and tolerance to multiple antibiotics and stresses in Escherichia coli.FEMS Microbiol Lett 303: 33–40. doi: 10.1111/j.1574-6968.2009.01857.x
|
| [64] |
Kwon O, Kotsakis A, Meganathan R (2000) Ubiquinone (coenzyme Q) biosynthesis in Escherichia coli: identification of the ubiF gene. FEMS Microbiol Lett 186: 157–161. doi: 10.1111/j.1574-6968.2000.tb09097.x
|
| [65] | Vázquez-Laslop N, Lee H, Neyfakh AA (2006) Increased persistence in Escherichia coli caused by controlled expression of toxins or other unrelated proteins.J Bacteriol 188: 3494–3497. |
| [66] |
Lee JH, Kim YG, Gwon G, et al. (2016) Halogenated indoles eradicate bacterial persister cells and biofilms. AMB Express 6: 123. doi: 10.1186/s13568-016-0297-6
|
| [67] |
Mina EG, Marques CNH (2016) Interaction of Staphylococcus aureus persister cells with the host when in a persister state and following awakening. Sci Reports 6: 31342. doi: 10.1038/srep31342
|
| [68] | Marques CNH, Morozov A, Planzos P, et al. (2014) The fatty acid signaling molecule cis-2-decenoic acid increases metabolic activity and reverts persister cells to an antimicrobial susceptible state. Appl Environ Microbiol 10: 6976–6991. |
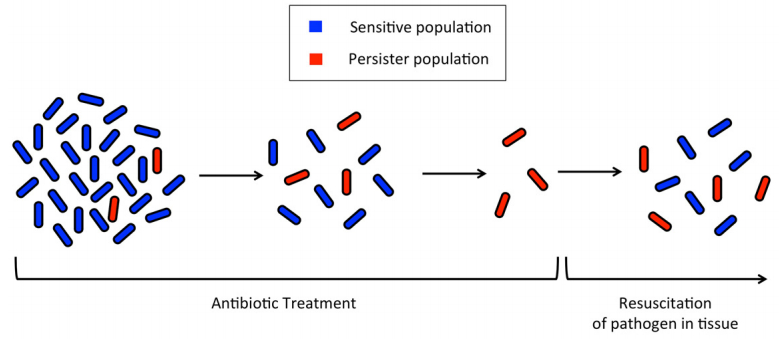
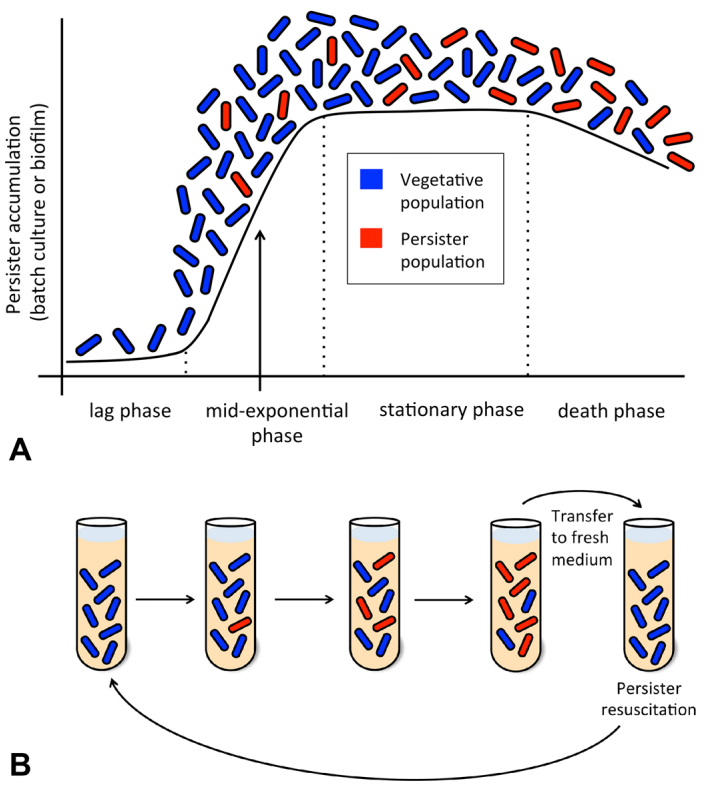
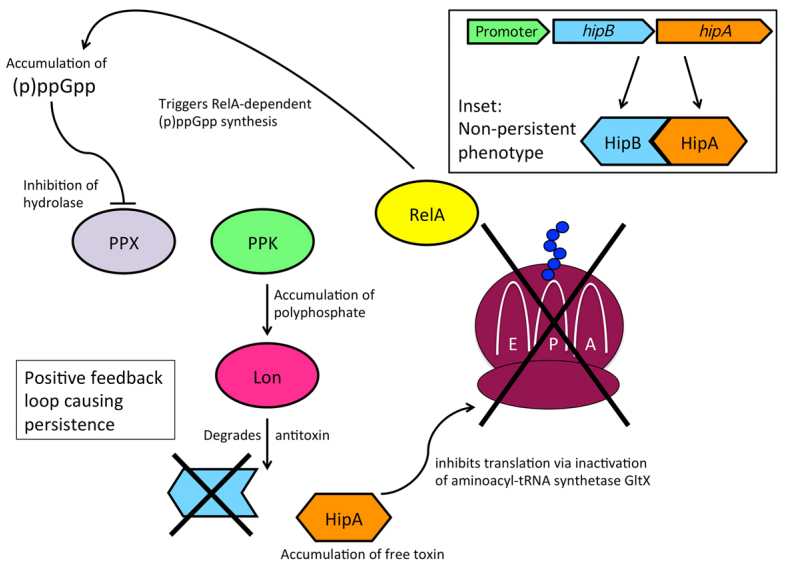
Figures(3) / Tables(1)

Tara L. Renbarger, Jennifer M. Baker, W. Matthew Sattley. Slow and steady wins the race: an examination of bacterial persistence[J]. AIMS Microbiology, 2017, 3(2): 171-185. doi: 10.3934/microbiol.2017.2.171







 DownLoad:
DownLoad: