Citation: Benjamin P Jones, Srdjan Saso, Timothy Bracewell-Milnes, Jen Barcroft, Jane Borley, Teodor Goroszeniuk, Kostas Lathouras, Joseph Yazbek, J Richard Smith. Laparoscopic uterosacral nerve block: A fertility preserving option in chronic pelvic pain[J]. AIMS Medical Science, 2019, 6(4): 260-267. doi: 10.3934/medsci.2019.4.260

| [1] | Donaldson L (2009) 150 years of the Annual Report of the Chief Medical Officer: On the state of public health 2008. London: Department of Health. |
| [2] | Ahangari A (2014) Prevalence of chronic pelvic pain among women: An updated review. Pain Physician 17: E141–147. |
| [3] |
van Wilgen CP, Keizer D (2012) The sensitization model to explain how chronic pain exists without tissue damage. Pain Manag Nurs 13: 60–65. doi: 10.1016/j.pmn.2010.03.001

|
| [4] |
Lamvu G (2011) Role of hysterectomy in the treatment of chronic pelvic pain. Obstet Gynecol 117: 1175–1178. doi: 10.1097/AOG.0b013e31821646e1
|
| [5] |
Mathias SD, Kuppermann M, Liberman RF, et al. (1996) Chronic pelvic pain: prevalence, health-related quality of life, and economic correlates. Obstet Gynecol 87: 321–327. doi: 10.1016/0029-7844(95)00458-0
|
| [6] | Royal Collge of Obstetricians & Gynaecologists (RCOG) (2015) Scientific Impact Paper No. 46. Therapies Targeting the Nervous System for Chronic Pelvic Pain Relief. RCOG: London |
| [7] |
Lee TT, Yang LC (2008) Pelvic denervation procedures: A current reappraisal. Int J Gynaecol Obstet 101: 304–308. doi: 10.1016/j.ijgo.2008.02.010
|
| [8] |
Huber SA, Northington GM, Karp DR (2015) Bowel and bladder dysfunction following surgery within the presacral space: an overview of neuroanatomy, function, and dysfunction. Int Urogynecol J 26: 941–946. doi: 10.1007/s00192-014-2572-x
|
| [9] |
Chen FP, Soong YK (1997) The efficacy and complications of laparoscopic presacral neurectomy in pelvic pain. Obstet Gynecol 90: 974–977. doi: 10.1016/S0029-7844(97)00484-5
|
| [10] | Lichten EM, Bombard J (1987) Surgical treatment of primary dysmenorrhea with laparoscopic uterine nerve ablation. J Reprod Med 32: 37–41. |
| [11] | Daniels JP, Middleton L, Xiong T, et al. (2010) International LUNA IPD Meta-analysis Collaborative Group. Individual patient data meta-analysis of randomized evidence to assess the effectiveness of laparoscopic uterosacral nerve ablation in chronic pelvic pain. Hum Reprod Update 16: 568–576. |
| [12] |
El-Din Shawki H (2011) The efficacy of laparoscopic uterosacral nerve ablation (LUNA) in the treatment of unexplained chronic pelvic pain: a randomized controlled trial. Gynecol Surg 8: 31–39. doi: 10.1007/s10397-010-0612-1
|
| [13] | Daniels J, Gray R, Hills RK, et al. (2009) LUNA Trial Collaboration. Laparoscopic uterosacral nerve ablation for alleviating chronic pelvic pain: A randomized controlled trial. JAMA 302: 955–961. |
| [14] | Jedrzejczak P, Sokalska A, Spaczynski RZ, et al. (2009) Effects of presacral neurectomy on pelvic pain in women with and without endometriosis. Ginekol Pol 80: 172–178. |
| [15] |
Rouholamin S, Jabalameli M, Mostafa A (2015) The effect of preemptive pudendal nerve block on pain after anterior and posterior vaginal repair. Adv Biomed Res 27: 153. doi: 10.4103/2277-9175.161580
|
| [16] | Chanrachakul B, Likittanasombut P, O-Prasertsawat P, et al. (2001) Lidocaine versus plain saline for pain relief in fractional curettage: A randomized controlled trial. Obstet Gynecol 98: 592–595. |
| [17] |
Naghshineh E, Shiari S, Jabalameli M (2015) Preventive effect of ilioinguinal nerve block on postoperative pain after cesarean section. Adv Biomed Res 4: 229. doi: 10.4103/2277-9175.166652
|
| [18] |
Binkert CA, Hirzel FC, Gutzeit A, et al. (2015) Superior hypogastric nerve block to reduce pain after uterine artery embolization: Advanced technique and comparison to epidural anesthesia. Cardiovasc Intervent Radiol 38: 1157–1161. doi: 10.1007/s00270-015-1118-z
|
| [19] |
Rapp H, Ledin Eriksson S, Smith P (2017) Superior hypogastric plexus block as a new method of pain relief after abdominal hysterectomy: Double-blind, randomised clinical trial of efficacy. BJOG 124: 270–276. doi: 10.1111/1471-0528.14119
|
| [20] |
Fujii M, Sagae S, Sato T, et al. (2002) Investigation of the localization of nerves in the uterosacral ligament: Determination of the optimal site for uterosacral nerve ablation. Gynecol Obstet Invest 54: discussion 16–7. doi: 10.1159/000066289
|
| [21] |
Matalliotakis IM, Katsikis IK, Panidis DK (2005) Adenomyosis: What is the impact on fertility? Curr Opin Obstet Gynecol 17: 261–264. doi: 10.1097/01.gco.0000169103.85128.c0
|
| [22] | Desrosiers JA, Faucher GL (1964) Uterosacral block: A new diagnostic procedure. Obstet Gynecol 23: 671–677. |
| [23] |
Rana MV, Candido KD, Raja O, et al. (2014) Celiac plexus block in the management of chronic abdominal pain. Curr Pain Headache Rep 18: 394. doi: 10.1007/s11916-013-0394-z
|
| [24] |
Soysal ME, Soysal S, Gurses E, et al. (2003) Laparoscopic presacral neurolysis for endometriosis-related pelvic pain. Hum Reprod 18: 588–592. doi: 10.1093/humrep/deg127
|
| [25] |
Byrd D, Mackey S (2008) Pulsed radiofrequency for chronic pain. Curr Pain Headache Rep 12: 37–41. doi: 10.1007/s11916-008-0008-3
|
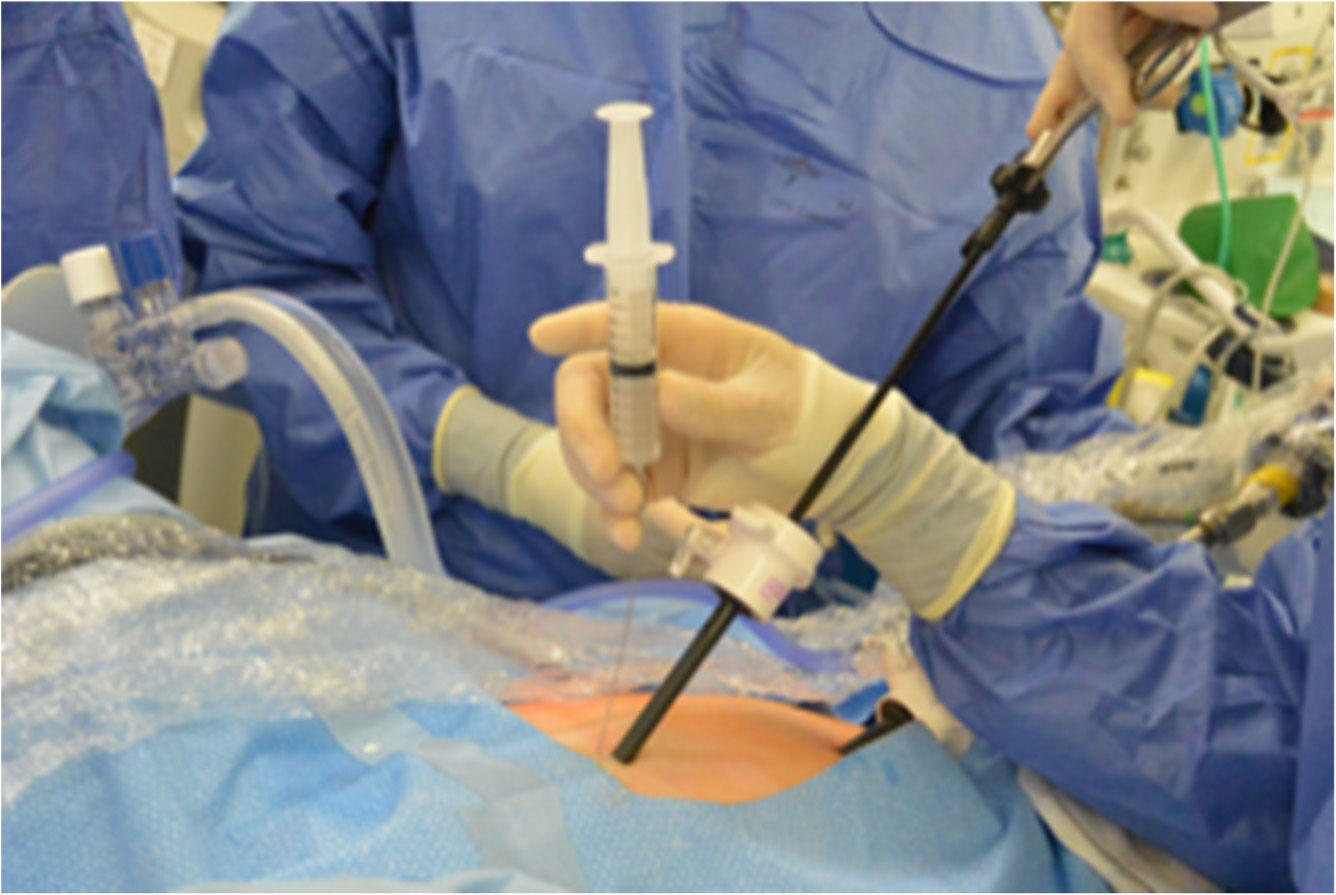
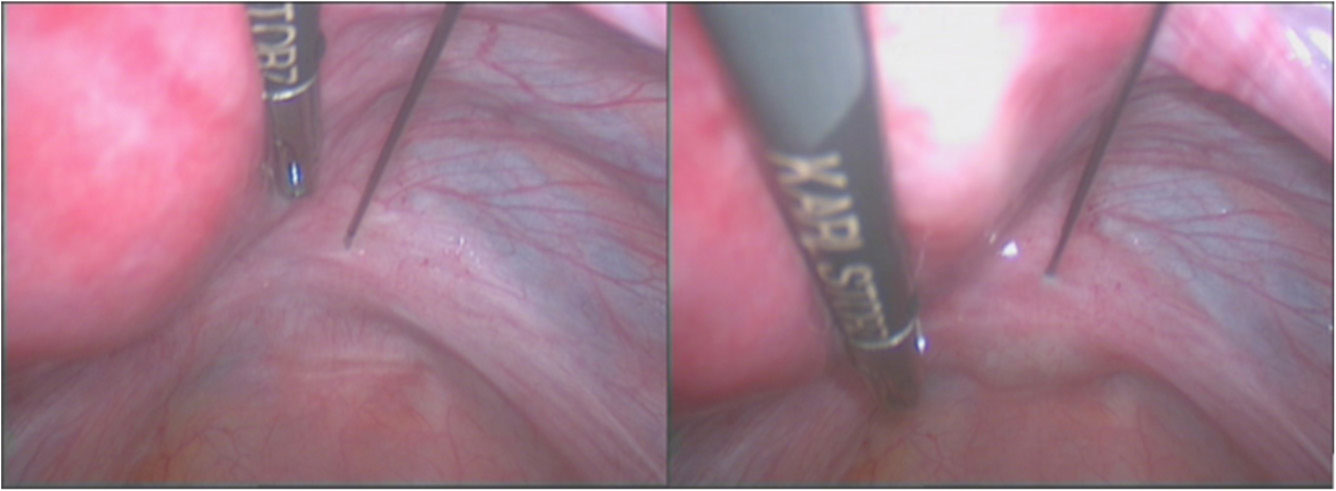
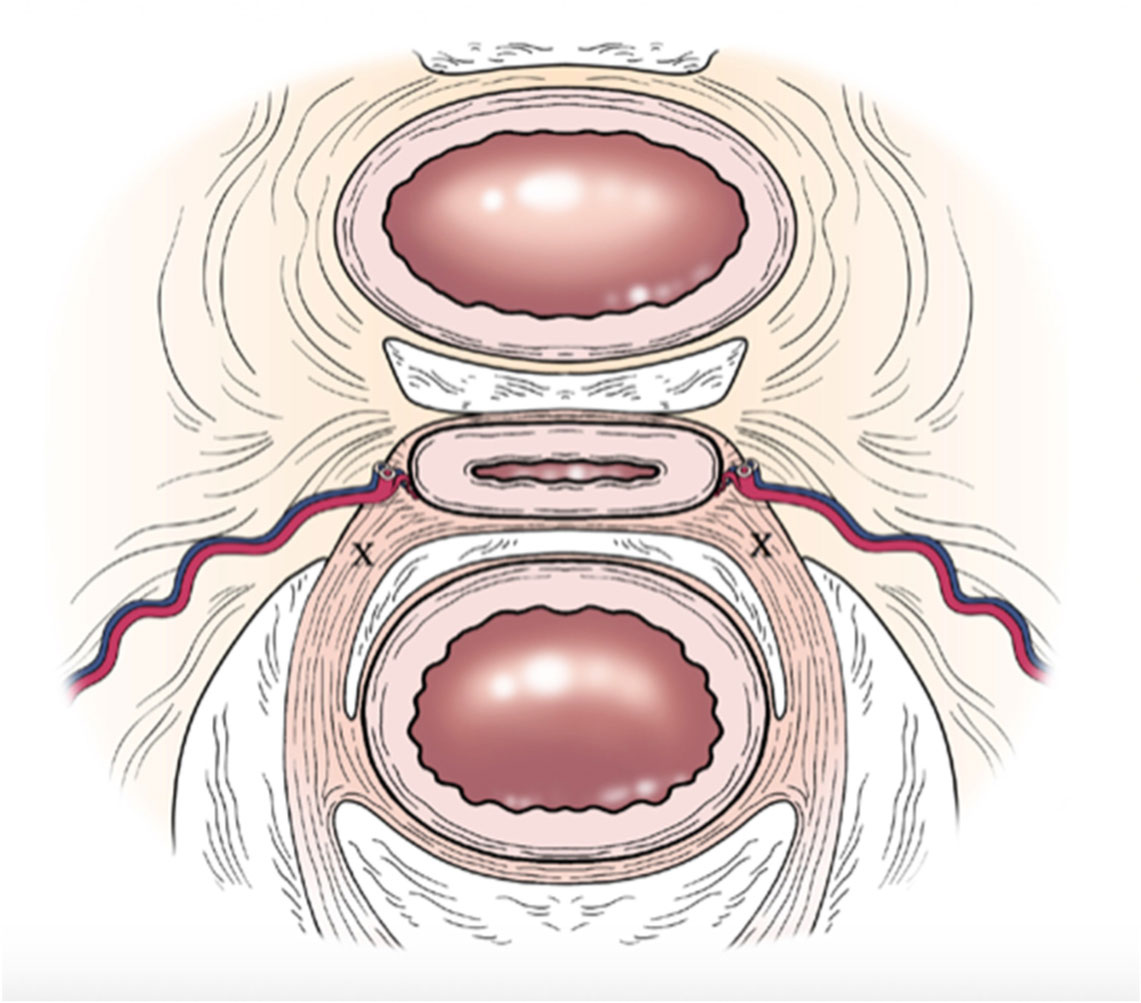
Figures(3)

Benjamin P Jones, Srdjan Saso, Timothy Bracewell-Milnes, Jen Barcroft, Jane Borley, Teodor Goroszeniuk, Kostas Lathouras, Joseph Yazbek, J Richard Smith. Laparoscopic uterosacral nerve block: A fertility preserving option in chronic pelvic pain[J]. AIMS Medical Science, 2019, 6(4): 260-267. doi: 10.3934/medsci.2019.4.260







 DownLoad:
DownLoad: