Citation: Ting Guo, Zhipeng Qiu. The effects of CTL immune response on HIV infection model with potent therapy, latently infected cells and cell-to-cell viral transmission[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6822-6841. doi: 10.3934/mbe.2019341

| [1] | A. S. Perelson, D. E. Kirschner and R. D. Boer, Dynamics of HIV infection of CD4+ T cells, Math. Biosci., 114 (1993), 81–125. |
| [2] | Y. Wang, J. Liu and L. Liu, Viral dynamics of an HIV model with latent infection incorporating antiretroviral therapy, Adv. Differ. Equations, 225 (2016). |
| [3] | WHO, 10 facts on HIV/AIDS, 2017. Available from: http://www.who.int/features/factfiles/hiv/zh/. |
| [4] | WHO, HIV/AIDS: Fact sheet, 2017. Available from: http://www.who.int/mediacentre/factsheets/fs360/en/. |
| [5] | A. Mojaver and H. Kheiri, Mathematical analysis of a class of HIV infection models of CD4+ T-cells with combined antiretroviral therapy, Appl. Math. Comput., 259 (2015), 258–270. |
| [6] | X. Wang, X. Song, S. Tang, et al., Dynamics of an HIV Model with Multiple Infection Stages and Treatment with Different Drug Classes, Bull. Math. Biol., 78 (2016), 322–349. |
| [7] | L. Rong and A. S. Perelson, Modeling HIV persistence, the latent reservoir, and viral blips, J. Theor. Biol., 260 (2009), 308–331. |
| [8] | J. M. Kitayimbwa, J. T. Mugisha and R. A. Saenz, The role of backward mutations on the within-host dynamics of HIV-1, J. Math. Biol., 67 (2013), 1111–1139. |
| [9] | S. Palmer, L. Josefsson and J. M. Coffin, HIV reservoirs and the possibility of a cure for HIV infection, J. Intern. Med., 270 (2011), 550–560. |
| [10] | L. Rong and A. S. Perelson, Modeling Latently Infected Cell Activation: Viral and Latent Reservoir Persistence, and Viral Blips in HIV-infected Patients on Potent Therapy, Plos Comput. Biol., 5 (2009). |
| [11] | F. Maldarelli, S. Palmer, M. S. King, et al., ART suppresses plasma HIV-1 RNA to a stable set point predicted by pretherapy viremia, Plos Pathog., 3 (2007). |
| [12] | H. S. Ariel, C. L. Lu, K. Florian, et al., Broadly Neutralizing Antibodies and Viral Inducers Decrease Rebound from HIV-1 Latent Reservoirs in Humanized Mice, Cell, 158 (2014), 989–999. |
| [13] | X. Wang, G. Mink, D. Lin, et al., Influence of raltegravir intensification on viral load and 2-LTR dynamics in HIV patients on suppressive antiretroviral therapy, J. Theor. Biol., 416 (2017), 16–27. |
| [14] | A. Bosque, K. A. Nilson, A. B. Macedo, et al., Benzotriazoles Reactivate Latent HIV-1 through Inactivation of STAT5 SUMOylation, Cell Rep., 18 (2017), 1324–1334. |
| [15] | S. Pankavich, The Effects of Latent Infection on the Dynamics of HIV, Differ. Equ. Dyn. Syst., 24 (2016), 281–303. |
| [16] | C. M. Pinto, Persistence of low levels of plasma viremia and of the latent reservoir in patients under ART: A fractional-order approach, Commun. Nonlinear Sci. Numer. Simulat., 43 (2017), 251–260. |
| [17] | D. C. Johnson and M. T. Huber, Directed egress of animal viruses promotes cell-to-cell spread, J. Virol., 76 (2002), 1–8. |
| [18] | D. Mazurov, A. Ilinskaya, G. Heidecker, et al., Quantitative comparison of HTLV-1 and HIV-1 cell-to-cell infection with new replication dependent vectors, Plos Path., 6 (2010). |
| [19] | H. Sato, J. Orenstein, D. Dimitrov, et al., Cell-to-cell spread of HIV-1 occurs within minutes and may not involve the participation of virus particles, Virology, 186 (1992), 712–724. |
| [20] | C. J. Duncan, R. A. Russell and Q. J. Sattentau, High multiplicity HIV-1 cell-to-cell transmission from macrophages to CD4+ T cells limits antiretroviral efficacy, AIDS, 27 (2013), 2201–2206. |
| [21] | J. Wang, J. Pang, T. Kuniya, et al., Global threshold dynamics in a five-dimensional virus model with cell-mediated, humoral immune responses and distributed delays, Appl. Math. Comput., 241 (2014), 298–316. |
| [22] | Y. Nakata, Global dynamics of a cell mediated immunity in viral infection models with distributed delays, J. Math. Anal. Appl., 375 (2010), 14-27. |
| [23] | Z. Yuan, Z. Ma and X. Tang, Global stability of a delayed HIV infection model with nonlinear incidence rate, Nonlinear Dynam., 68 (2012), 207–214. |
| [24] | Z. Yuan and X. Zou, Global threshold dynamics in an HIV virus model with nonlinear infection rate and distributed invasion and production delays, Math. Biosci. Eng., 10 (2013), 483–498. |
| [25] | R. Arnaout, M. Nowak and D. Wodarz, HIV-1 dynamics revisited: biphasic decay by cytotoxic lymphocyte killing?, Proc. R. Soc. London, 265 (2000), 1347–1354. |
| [26] | J. M. Conway and A. S. Perelson, Post-treatment control of HIV infection, Proc. Natl. Acad. Sci. B, 112 (2015), 5467–5472. |
| [27] | Y. Wang, Y. Zhou, F.Brauer, et al., Viraldynamics model with CTL immuneresponse incorporating antiretroviral therapy, J. Math. Biol., 67 (2013), 901-934. |
| [28] | H. Pourbashash, S. S. Pilyugin, C. McCluskey, et al., Global analysis of within host virus models with cell-to-cell viral transmission, Discrete Contin. Dyn. Syst. Ser. B, 19 (2014), 3341–3357. |
| [29] | B. Song, J. Lou and Q. Wen, Modelling two different therapy strategies for drug T-20 on HIV-1 patients, J. Appl. Math. Mech., 32 (2011), 419–436. |
| [30] | D. S. Callaway and A. S. Perelson, HIV-1 infection and low steady state viral loads, Bull. Math. Biol., 64 (2002), 29–64. |
| [31] | K. Allali, J. Danane and Y. Kuang, Global analysis for an HIV infection model with CTL immune response and infected cells in eclipse phase, Appl. Sci., 7 (2017), 861. |
| [32] | L. Rong, Z. Feng and A. S. Perelson, Emergence of HIV-1 Drug Resistance During Antiretroviral Treatment, Bull. Math. Biol., 69 (2007), 2027–2060. |
| [33] | H. Zhu, Y. Luo and M. Chen, Stability and Hopf bifurcation of a HIV infection model with CTL- response delay, Comput. Math. Appl., 62 (2011), 3091–3102. |
| [34] | X. Wang, A. M. Elaiw and X. Song, Global properties of a delayed HIV infection model with CTL immune response, Appl. Math. Comput., 218 (2012), 9405–9414. |
| [35] | B. M. Adams, H. T. Banks, M. Davidian, et al., HIV dynamics: Modeling, data analysis, and optimal treatment protocols, J. Comput. Appl. Math., 184 (2005), 10–49. |
| [36] | L. Rong and A. S. Perelson, Asymmetric division of activated latently infected cells may explain the decay kinetics of the HIV-1 latent reservoir and intermittent viral blips, Math. Biosci., 217 (2009), 77–87. |
| [37] | P. Driessche and P. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. |
| [38] | J. P. LaSalle, The stability of dynamical systems, Philadelphia, 1976. |
| [39] | X. Tian and R. Xu, Global stability and Hopf bifurcation of an HIV-1 infection model with saturation incidence and delayed CTL immune response, Appl. Math. Comput., 237 (2014), 146–154. |
| [40] | M. Louie, C. Hogan, M. D. Mascio, et al., Determining the relative efficacy of highly active antiretroviral therapy, J. Infect. Dis., 187 (2003), 896–900. |
| [41] | M. A. Nowak and C. R. Bangham, Population dynamics of immune responses to persistent viruses, Science, 272 (1996), 74–79. |
| [42] | A. S. Perelson, P. Essunger, Y. Cao, et al., Decay characteristics of HIV-1-infected compartments during combination therapy, Nature, 387 (1997), 188–191. |
| [43] | J. Wang, M. Guo, X. Liu, et al., Threshold dynamics of HIV-1 virus model with cell-to-cell transmission, cell-mediated immune responses and distributed delay, Appl. Math. Comput., 291 (2016), 149–161. |
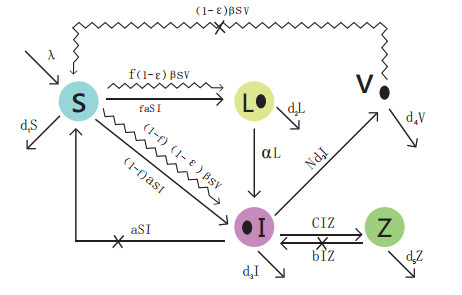
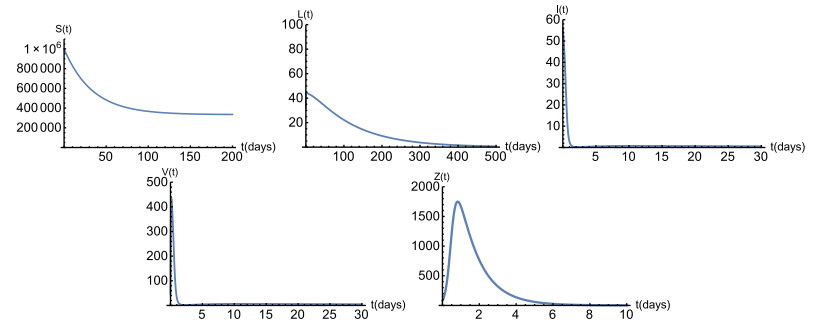
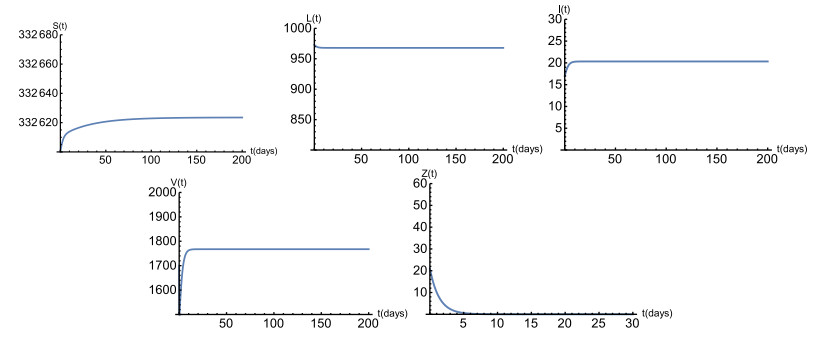
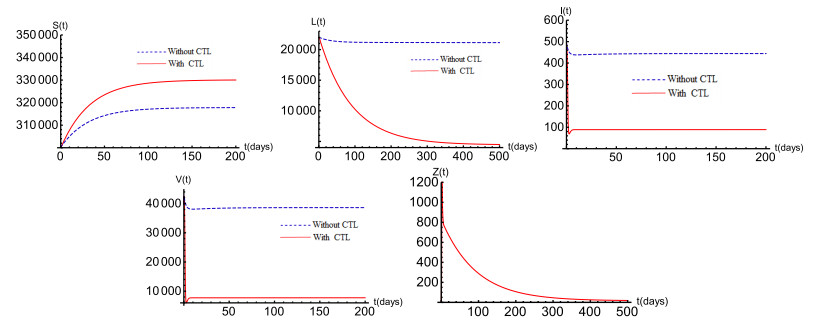
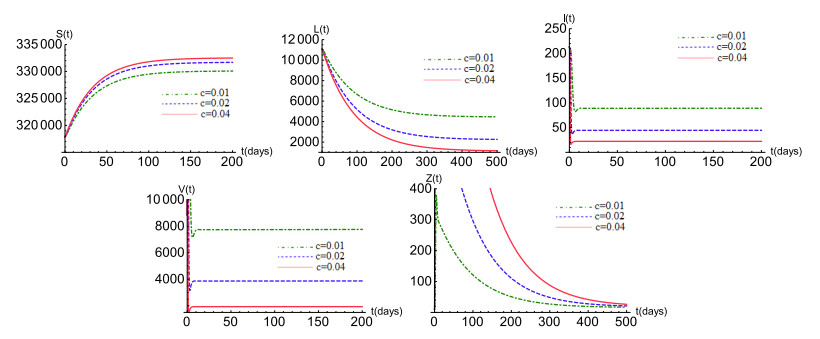
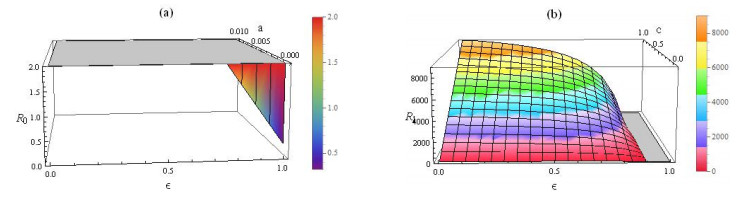
Figures(6) / Tables(1)

Ting Guo, Zhipeng Qiu. The effects of CTL immune response on HIV infection model with potent therapy, latently infected cells and cell-to-cell viral transmission[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6822-6841. doi: 10.3934/mbe.2019341







 DownLoad:
DownLoad: