Citation: Ying Jiang, Min Luo, Jianfeng Jiao, Ruiqi Wang. Study on cell apoptosis based on bifurcation and sensitivity analysis[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 3235-3250. doi: 10.3934/mbe.2019161

| [1] | D. Hanahan and R. A. Weinberg, The hallmarks of cancer, Cell,100(2000), 57–70. |
| [2] | D. W. Nicholson, From bench to clinic with apoptosis-based therapeutic agents, Nature, 407 (2000), 810–816. |
| [3] | F. Nezhat, C. Cohen, J. Rahaman, et al., Comparative immunohistochemical studies of bcl-2 and p53proteinsinbenignandmalignantovarianendometrioticcysts, Cancer,94(2002),2935–2940. |
| [4] | K. Nasu, M. Nishida, Y. Kawano, et al., Aberrant expression of apoptosis-related molecules in endometriosis a possible mechanism underlying the pathogenesis of endometriosis, Reprod. Sci., 18(2011), 206–218. |
| [5] | J. M. Lee and A. Bernstein, Apoptosis, cancer and the p53 tumour suppressor gene, Cancer Metastasis Rev.,14(1995), 149–161. |
| [6] | A. Ashkenazi, Targeting the extrinsic apoptosis pathway in cancer, Cytokine Growth Factor Rev., 19(2008), 325–331. |
| [7] | T. L. Zhang, P. Brazhnik and J. J. Tyson, Computational analysis of dynamical responses to the intrinsic pathway of programmed cell death, Biophys. J.,97(2009), 415–434. |
| [8] | J. Chen, H. Yue and Q. Ouyang, Correlation between oncogenic mutations and parameter sensitivity of the apoptosis pathway model, PLoS Comput. Biol.,10(2014), e1003451. |
| [9] | L. Zhao, T. L. Sun, J. Pei, et al., Mutation-induced protein interaction kinetics changes affect apoptotic network dynamic properties and facilitate oncogenesis, Proc. Natl. Acad. Sci. U. S. A.,112(2015), E4046–E4054. |
| [10] | X. L. Chen, J. Chen, B. Shao, et al., Relationship between cancer mutations and parameter sensitivity in Rb pathway, J. Theor. Biol.,404(2016), 120–125. |
| [11] | J. E. Ferrell, J. R. Pomerening, S. Y. Kim, et al., Simple, realistic models of complex biological processes: positive feedback and bistability in a cell fate switch and a cell cycle oscillator, FEBS Lett.,583(2009), 3999–4005. |
| [12] | M. Y. Lu, M. K. Jolly, H. Levine, et al., MicroRNA-based regulation of epithelial-hybridmesenchymal fate determination, Proc. Natl. Acad. Sci. U. S. A.,110(2013), 18144–18149. |
| [13] | Y. S. Lee, O. Z. Liu, H. S. Hwang, et al., Parameter sensitivity analysis of stochastic models provides insights into cardiac calcium sparks, Biophys. J.,104(2013), 1142–1150. |
| [14] | H. Yue, M. Brown, J. Knowles, et al., Insights into the behaviour of systems biology models from dynamicsensitivityandidentifiabilityanalysis: acasestudyofanNF-kappaBsignallingpathway, Mol. Biosyst.,2(2006), 640–649. |
| [15] | S. Desagher, A. O. Sand, A. Nichols, et al., Bid-induced conformational change of bax is responsible for mitochondrial cytochrome c release during apoptosis, J. Cell Biol.,144(1999), 891–901. |
| [16] | R. Eskes, S. Desagher, B. Antonsson, et al., Bid induces the oligomerization and insertion of Bax into the outer mitochondrial membrane, Mol. Cell. Biol.,20(2000), 929–935. |
| [17] | M. Marani, T. Tenev, D. Hancock, et al., Identification of novel isoforms of the BH3 domain protein bim which directly activate Bax to trigger apoptosis, Mol. Cell. Biol., 22 (2002), 3577– 3589. |
| [18] | J. G. Albeck, J. M. Burke, B. B. Aldridge, et al., Quantitative analysis of pathways controlling extrinsic apoptosis in single cells, Mol. Cell,30(2008), 11–25. |
| [19] | J. C. Goldstein, N. J. Waterhouse, P. Juin, et al., The coordinate release of cytochrome c during apoptosis is rapid,complete and kinetically invariant, Nat. Cell Biol.,2(2000), 156–162. |
| [20] | J. S. Yuan, Y. Deng, Y. Y. Zhang, et al., Bmp4 inhibits goose granulosa cell apoptosis via PI3K/AKT/Caspase-9 signaling pathway, Anim. Reprod. Sci.,200(2018), 86–95. |
| [21] | F. Wang, Z. L. Wei, X. R. Sun, et al., Apoptosis inducing factor is involved in stretch-induced apoptosis of myoblast via a caspase-9 independent pathway, J. Cell. Biochem., 118 (2017), 829– 838. |
| [22] | R. Reinhart, L. Rohner, S. Wicki, et al., BH3 mimetics efficiently induce apoptosis in mouse basophils and mast cells, Cell Death Differ.,25(2018), 204–216. |
| [23] | L.Li,Q.Tao,H.Jin,etal., ThetumorsuppressorUCHL1formsacomplexwithp53/MDM2/ARF to promote p53 signaling and is frequently silenced in nasopharyngeal carcinoma, Clin. Cancer Res.,16(2010), 2949–2958. |
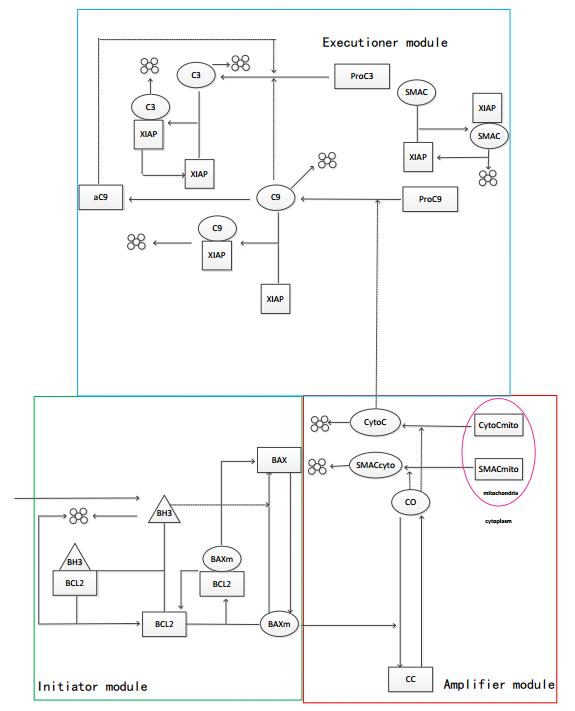
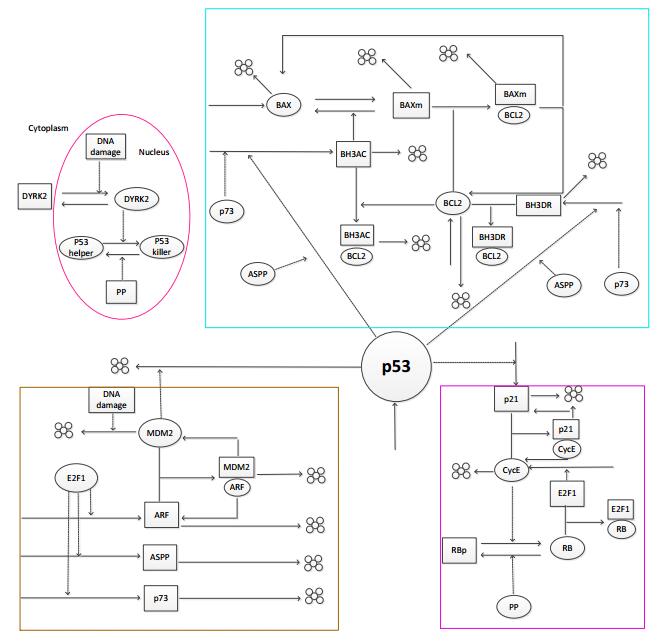
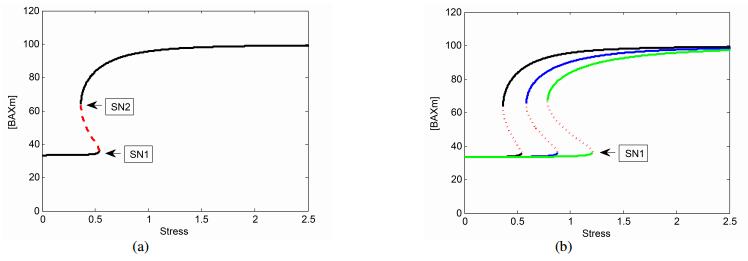
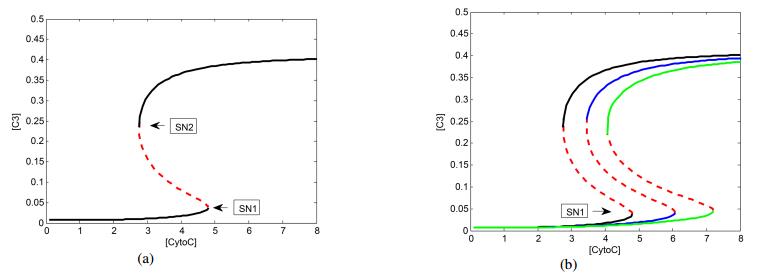
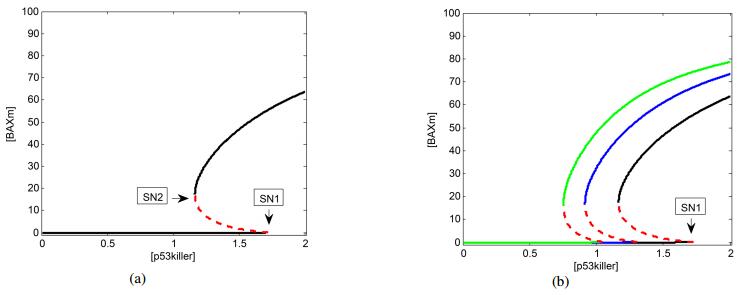
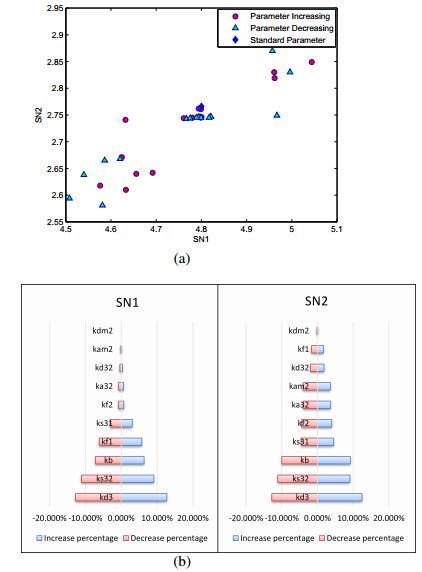
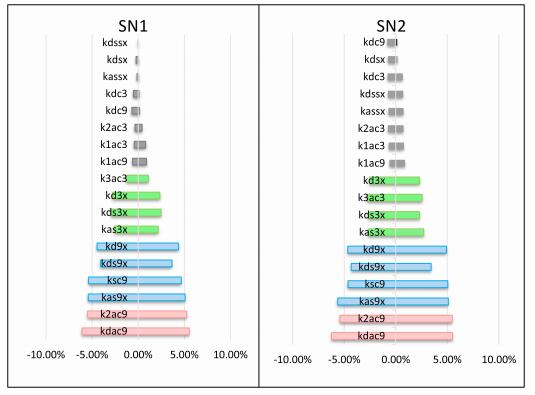
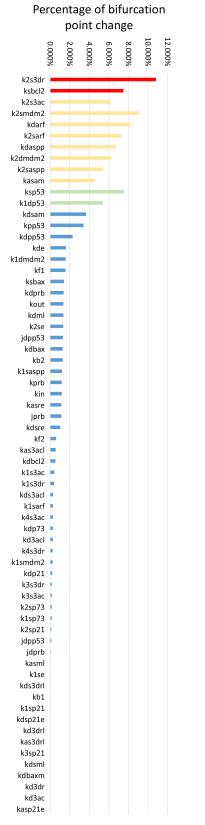
Figures(8)

Ying Jiang, Min Luo, Jianfeng Jiao, Ruiqi Wang. Study on cell apoptosis based on bifurcation and sensitivity analysis[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 3235-3250. doi: 10.3934/mbe.2019161







 DownLoad:
DownLoad: