Citation: Stephen A. Gourley, Xiulan Lai, Junping Shi, Wendi Wang, Yanyu Xiao, Xingfu Zou. Role of white-tailed deer in geographic spread of the black-legged tick Ixodes scapularis : Analysis of a spatially nonlocal model[J]. Mathematical Biosciences and Engineering, 2018, 15(4): 1033-1054. doi: 10.3934/mbe.2018046

| [1] | [ R. M. Bacon,K. J. Kugeler,K. S. Griffith,P. S. Mead, Lyme disease -United States, 2003-2005, Journal of the American Medical Association, 298 (2007): 278-279. |
| [2] | [ A. G. Barbour,D. Fish, The biological and social phenomenon of Lyme disease, Science, 260 (1993): 1610-1616. |
| [3] | [ R. J. Brinkerhoff,C. M. Folsom-O'Keefe,K. Tsao,M. A. Diuk-Wasser, Do birds affect Lyme disease risk? Range expansion of the vector-borne pathogen Borrelia burgdorferi, Front. Ecol. Environ, 9 (2011): 103-110. |
| [4] | [ S. G. Caraco,S. Glavanakov,G. Chen,J. E. Flaherty,T. K. Ohsumi,B. K. Szymanski, Stage-structured infection transmission and a spatial epidemic: a model for Lyme disease, Am. Nat., 160 (2002): 348-359. |
| [5] | [ M. R. Cortinas,U. Kitron, County-level surveillance of white-tailed deer infestation by Ixodes scapularis and Dermacentor albipictus (Acari: Ixodidae) along the Illinois River, J. Med. Entomol., 43 (2006): 810-819. |
| [6] | [ D. T. Dennis,T. S. Nekomoto,J. C. Victor,W. S. Paul,J. Piesman, Reported distribution of Ixodes scapularis and Ixodes pacificus (Acari: Ixodidae) in the United States, J. Med. Entomol., 35 (1998): 629-638. |
| [7] | [ G. Fan,H. R. Thieme,H. Zhu, Delay differential systems for tick population dynamics, J. Math. Biol., 71 (2015): 1017-1048. |
| [8] | [ S. A. Gourley,S. Ruan, A delay equation model for oviposition habitat selection by mosquitoes, J. Math. Biol., 65 (2012): 1125-1148. |
| [9] | [ B. H. Hahn,C. S. Jarnevich,A. J. Monaghan,R. J. Eisen, Modeling the Geographic Distribution of Ixodes scapularis and Ixodes pacificus (Acari: Ixodidae) in the Contiguous United States, Journal of Medical Entomology, 53 (2016): 1176-1191. |
| [10] | [ S. Hamer,G. Hickling,E. Walker,J. I. Tsao, Invasion of the Lyme disease vector Ixodes scapularis: Implications for Borrelia burgdorferi endemicity, EcoHealth, 7 (2010): 47-63. |
| [11] | [ X. Lai and X. Zou, Spreading speed and minimal traveling wave speed in a spatially nonlocal model for the population of blacklegged tick Ixodes scapularis, in preparation. |
| [12] | [ J. Li,X. Zou, Modeling spatial spread of infectious diseases with a fixed latent period in a spatially continuous domain, Bull. Math. Biol., 71 (2009): 2048-2079. |
| [13] | [ D. Liang,J. W.-H. So,F. Zhang,X. Zou, Population dynamic models with nonlocal delay on bounded fields and their numeric computations, Diff. Eqns. Dynam. Syst., 11 (2003): 117-139. |
| [14] | [ K. Liu,Y. Lou,J. Wu, Analysis of an age structured model for tick populations subject to seasonal effects, J. Diff. Eqns., 263 (2017): 2078-2112. |
| [15] | [ N. K. Madhav,J. S. Brownstein,J. I. Tsao,D. Fish, A dispersal model for the range expansion of blacklegged tick (Acari: Ixodidae), J. Med. Entomol., 41 (2004): 842-852. |
| [16] | [ M. G. Neubert,I. M. Parker, Projecting rates of spread for invasive species, Risk Analysis, 24 (2004): 817-831. |
| [17] | [ N. H. Ogden,M. Bigras-Poulin,C. J. O'Callaghan,I. K. Barker,L. R. Lindsay,A. Maarouf,K. E. Smoyer-Tomic,D. Waltner-Toews,D. Charron, A dynamic population model to investigate effects of climate on geographic range and seasonality of the tick Ixodes scapularis, Int J. Parasitol., 35 (2005): 375-389. |
| [18] | [ N. H. Ogden,L. R. Lindsay,K. Hanincova,I. K. Barker,M. Bigras-Poulin,D. F. Charron,A. Heagy,C. M. Francis,C. J. O'Callaghan,I. Schwartz,R. A. Thompson, Role of migratory birds in introduction and range expansion of Ixodes scapularis ticks and of Borrelia burgdorferi and Anaplasma phagocytophilum in Canada, Applied and Environmental Microbiology, 74 (2008): 1780-1790. |
| [19] | [ J. W.-H. So,J. Wu,X. Zou, A reaction diffusion model for a single species with age structure -I. Traveling wave fronts on unbounded domains, Proc. Royal Soc. London. A, 457 (2001): 1841-1853. |
| [20] | [ H. R. Thieme, Spectral bound and reproduction number for infinite-dimensional population structure and time heterogeneity, SIAM J. Appl. Math., 70 (2009): 188-211. |
| [21] | [ J. Van Buskirk,R. S. Ostfeld, Controlling Lyme disease by modifying the density and species composition of tick hosts, Ecological Applications, 5 (1995): 1133-1140. |
| [22] | [ H. F. Weinberger,M. A. Lewis,B. Li, Analysis of linear determinacy for spread in cooperative models, J. Math. Biol., 45 (2002): 183-218. |
| [23] | [ X. Wu,G. Röst,X. Zou, Impact of spring bird migration on the range expansion of Ixodes scapularis tick population, Bull. Math. Biol., 78 (2016): 138-168. |
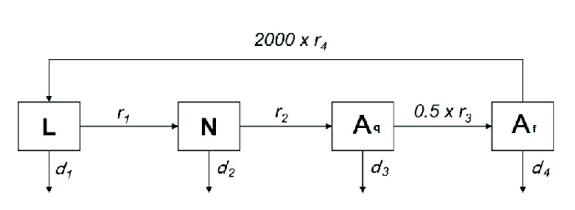
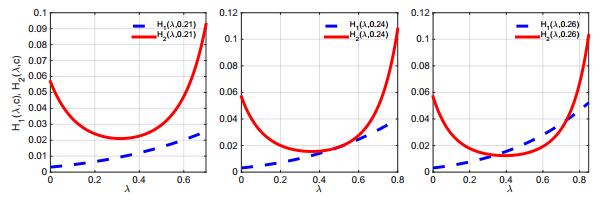
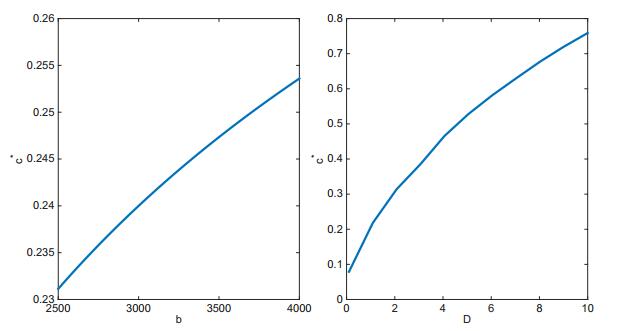
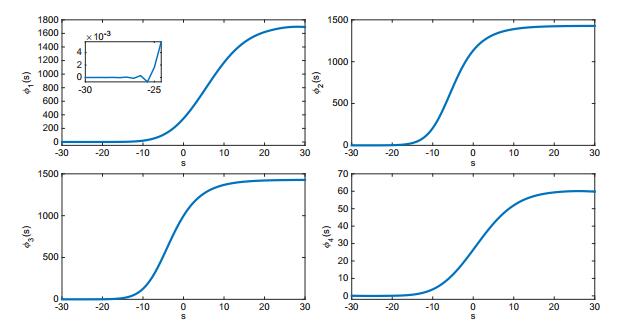
Figures(7) / Tables(1)

Stephen A. Gourley, Xiulan Lai, Junping Shi, Wendi Wang, Yanyu Xiao, Xingfu Zou. Role of white-tailed deer in geographic spread of the black-legged tick Ixodes scapularis : Analysis of a spatially nonlocal model[J]. Mathematical Biosciences and Engineering, 2018, 15(4): 1033-1054. doi: 10.3934/mbe.2018046







 DownLoad:
DownLoad: