Citation: Bruno Buonomo, Giuseppe Carbone, Alberto d'Onofrio. Effect of seasonality on the dynamics of an imitation-based vaccination model with public health intervention[J]. Mathematical Biosciences and Engineering, 2018, 15(1): 299-321. doi: 10.3934/mbe.2018013

| [1] | [ R. M. Anderson, The impact of vaccination on the epidemiology of infectious diseases, in The Vaccine Book, B. R. Bloom and P. -H. Lambert (eds. ) The Vaccine Book (Second Edition). Elsevier B. V. , Amsterdam, (2006), 3-31. |
| [2] | [ R. M. Anderson,R. M. May, null, Infectious Diseases of Humans. Dynamics and Control, Oxford University Press, Oxford, 1991. |
| [3] | [ N. Bacaër, Approximation of the basic reproduction number $R_0$ for vector-borne diseases with a periodic vector population, Bull. Math. Biol., 69 (2007): 1067-1091. |
| [4] | [ C. T. Bauch, Imitation dynamics predict vaccinating behaviour, Proc. R. Soc. B, 272 (2005): 1669-1675. |
| [5] | [ B. R. Bloom and P. -H. Lambert (eds. ), The Vaccine Book (Second Edition), Elsevier B. V. , Amsterdam, 2006. |
| [6] | [ F. Brauer, P. van den Driessche and J. Wu (eds. ), Mathematical Epidemiology, Lecture Notes in Mathematics. Mathematical biosciences subseries, vol. 1945, Springer, Berlin, 2008. |
| [7] | [ B. Buonomo,A. d'Onofrio,D. Lacitignola, Modeling of pseudo-rational exemption to vaccination for SEIR diseases, J. Math. Anal. Appl., 404 (2013): 385-398. |
| [8] | [ B. Buonomo, A. d'Onofrio and P. Manfredi, Public Health Intervention to shape voluntary vaccination: Continuous and piecewise optimal control, Submitted. |
| [9] | [ V. Capasso, null, Mathematical Structure of Epidemic Models, 2nd edition, Springer, 2008. |
| [10] | [ L. Cesari, Asymptotic Behavior and Stability Problems in Ordinary Differential Equations, Ergebnisse der Mathematik und ihrer Grenzgebiete, Band 16, Springer-Verlag, New York-Heidelberg, 1971. |
| [11] | [ C. Chicone, Ordinary Differential Equations with Applications, Texts in Applied Mathematics, 34, Springer, New York, 2006. |
| [12] | [ M. Choisy,J.-F. Guegan,P. Rohani, Dynamics of infectious diseases and pulse vaccination: Teasing apart the embedded resonance effects, Physica D, 223 (2006): 26-35. |
| [13] | [ W. Coppel, Stability and Asymptotic Behavior of Differential Equations, Heath, Boston, 1965. |
| [14] | [ O. Diekmann,J. A. P. Heesterbeek,J. A. J. Metz, On the definition and the computation of the basic reproduction ratio $R_0$ in models for infectious diseases in heterogeneous populations, J. Math. Biol., 28 (1990): 365-382. |
| [15] | [ A. d'Onofrio, Stability properties of pulses vaccination strategy in SEIR epidemic model, Math. Biosci., 179 (2002): 57-72. |
| [16] | [ A. d'Onofrio and P. Manfredi, Impact of Human Behaviour on the Spread of Infectious Diseases: a review of some evidences and models, Submitted. |
| [17] | [ A. d'Onofrio,P. Manfredi,P. Poletti, The impact of vaccine side effects on the natural history of immunization programmes: An imitation-game approach, J. Theor. Biol., 273 (2011): 63-71. |
| [18] | [ A. d'Onofrio,P. Manfredi,P. Poletti, The interplay of public intervention and private choices in determining the outcome of vaccination programmes, PLoS ONE, 7 (2012): e45653. |
| [19] | [ A. d'Onofrio,P. Manfredi,E. Salinelli, Vaccinating behaviour, information and the dynamics of SIR vaccine preventable diseases, Theor. Popul. Biol., 71 (2007): 301-317. |
| [20] | [ A. d'Onofrio,P. Manfredi,E. Salinelli, Fatal SIR diseases and rational exemption to vaccination, Math. Med. Biol., 25 (2008): 337-357. |
| [21] | [ D. Elliman,H. Bedford, MMR: Where are we now?, Archives of Disease in Childhood, 92 (2007): 1055-1057. |
| [22] | [ B. F. Finkenstadt,B. T. Grenfell, Time series modelling of childhood diseases: A dynamical systems approach, Appl. Stat., 49 (2000): 187-205. |
| [23] | [ N. C. Grassly,C. Fraser, Seasonal infectious disease epidemiology, Proc. Royal Soc. B, 273 (2006): 2541-2550. |
| [24] | [ M. W. Hirsch and H. Smith, Monotone dynamical systems, in Handbook of Differential Equations: Ordinary Differential Equations, vol. Ⅱ, Elsevier B. V. , Amsterdam, 2 (2005), 239-357. |
| [25] | [ IMI. Innovative medicine initiative. First Innovative Medicines Initiative Ebola projects get underway, Available from: http://www.imi.europa.eu/content/ebola-project-launch (accessed September 2016). |
| [26] | [ A. Kata, A postmodern Pandora's box: Anti-vaccination misinformation on the Internet, Vaccine, 28 (2010): 1709-1716. |
| [27] | [ A. Kata, Anti-vaccine activists, Web 2.0, and the postmodern paradigm-An overview of tactics and tropes used online by the anti-vaccination movement, Vaccine, 30 (2012): 3778-3789. |
| [28] | [ M. J. Keeling,P. Rohani, null, Modeling Infectious Diseases in Humans and Animals, Princeton University Press, Princeton, NJ, 2008. |
| [29] | [ W. P. London,J. A. Yorke, Recurrent outbreaks of measles, chickenpox and mumps. I. Seasonal variation in contact rates, Am. J. Epidemiol., 98 (1973): 453-468. |
| [30] | [ P. Manfredi and A. d'Onofrio (eds), Modeling the Interplay Between Human Behavior and the Spread of Infectious Diseases, Springer, New York, 2013. |
| [31] | [ M. Martcheva, An Introduction to Mathematical Epidemiology, Texts in Applied Mathematics, 61, Springer, New York, 2015. |
| [32] | [ E. Miller,N. J. Gay, Epidemiological determinants of pertussis, Dev. Biol. Stand., 89 (1997): 15-23. |
| [33] | [ J. D. Murray, Mathematical Biology. I. An Introduction. Third Edition, Interdisciplinary Applied Mathematics, 17. Springer-Verlag, New York, 2002. |
| [34] | [ Y. Nakata,T. Kuniya, Global dynamics of a class of SEIRS epidemic models in a periodic environment, J. Math. Anal. Appl., 325 (2010): 230-237. |
| [35] | [ G. Napier,D. Lee,C. Robertson,A. Lawson,K. G. Pollock, A model to estimate the impact of changes in MMR vaccine uptake on inequalities in measles susceptibility in Scotland, Stat. Methods Med. Res., 25 (2016): 1185-1200. |
| [36] | [ L. F. Olsen,W. M. Schaffer, Chaos versus noisy periodicity: Alternative hypotheses for childhood epidemics, Science, 249 (1990): 499-504. |
| [37] | [ T. Philipson, Private Vaccination and Public Health an empirical examination for U.S. Measles, J. Hum. Res., 31 (1996): 611-630. |
| [38] | [ I. B. Schwartz,H. L. Smith, Infinite subharmonic bifurcation in an SEIR epidemic model, J. Math. Biol., 18 (1983): 233-253. |
| [39] | [ H. E. Soper, The interpretation of periodicity in disease prevalence, J. R. Stat. Soc., 92 (1929): 34-73. |
| [40] | [ P. van den Driessche,J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002): 29-48. |
| [41] | [ A. van Lier,J. van de Kassteele,P. de Hoogh,I. Drijfhout,H. de Melker, Vaccine uptake determinants in The Netherlands, Eur. J. Public Health, 24 (2013): 304-309. |
| [42] | [ G. S. Wallace, Why Measles Matters, 2014, Available from: http://stacks.cdc.gov/view/cdc/27488/cdc_27488_DS1.pdf (accessed November 2016). |
| [43] | [ Z. Wang,C. T. Bauch,S. Bhattacharyya,A. d'Onofrio,P. Manfredi,M. Perc,N. Perra,M. Salathé,D. Zhao, Statistical physics of vaccination, Physics Reports, 664 (2016): 1-113. |
| [44] | [ W. Wang,X. Q. Zhao, Threshold dynamics for compartmental epidemic models in periodic environments, J. Dyn. Diff. Eq., 20 (2008): 699-717. |
| [45] | [ WHO, Vaccine safety basics, Available from: http://vaccine-safety-training.org/adverse-events-classification.html (accessed November 2016). |
| [46] | [ R. M. Wolfe,L. K. Sharp, Anti-vaccinationists past and present, Brit. Med. J., 325 (2002): p430. |
| [47] | [ F. Zhang,X. Q. Zhao, A periodic epidemic model in a patchy environment, J. Math. Anal. Appl., 325 (2007): 496-516. |
| [48] | [ X. Q. Zhao, Dynamical Systems in Population Biology, CMS Books Math. , vol. 16, Springer-Verlag, New York, 2003. |
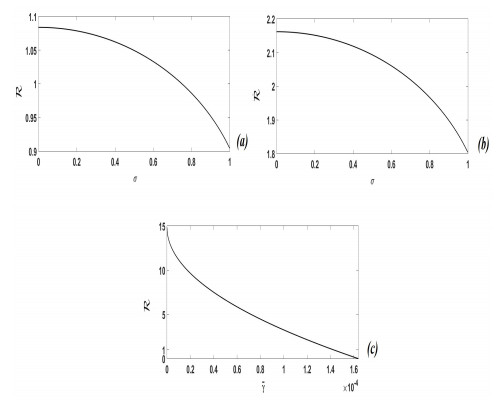
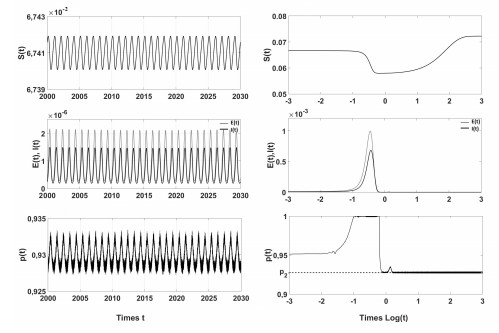
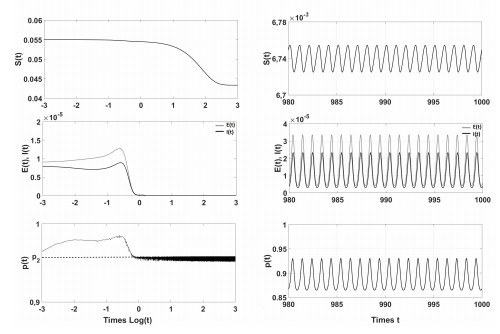
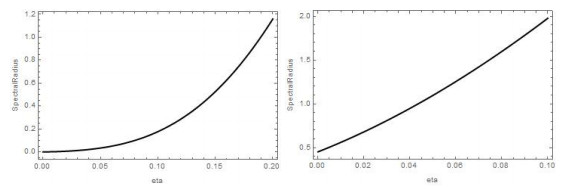
Figures(4)

Bruno Buonomo, Giuseppe Carbone, Alberto d'Onofrio. Effect of seasonality on the dynamics of an imitation-based vaccination model with public health intervention[J]. Mathematical Biosciences and Engineering, 2018, 15(1): 299-321. doi: 10.3934/mbe.2018013







 DownLoad:
DownLoad: