Citation: Stephen Tully, Monica-Gabriela Cojocaru, Chris T. Bauch. Multiplayer games and HIV transmission via casual encounters[J]. Mathematical Biosciences and Engineering, 2017, 14(2): 359-376. doi: 10.3934/mbe.2017023

| [1] | [ K. J. Arrow,G. Debreu, Existence of an equilibrium for a competitive economy, Econometrica, 22 (1954): 265-290. |
| [2] | [ J. P. Aubin and A. Cellina, Differential Inclusions: Set-Valued Maps and Viability Theory, Springer-Verlag New York, 1984. |
| [3] | [ R. M. Axelrod, The complexity of cooperation: Agent-based models of competition and collaboration, Princeton University Press, 1997. |
| [4] | [ T. Basar and G. J. Olsder, Dynamic Noncooperative Game Theory, SIAM, 1995. |
| [5] | [ A. Bensoussan, Points de Nash dans le cas de fonctionnelles quadratiques et jeux differentiels lineaires a N personnes, SIAM J. Control, 12 (1974): 460-499. |
| [6] | [ Center for Disease Control and Prevention, Diagnoses of HIV Infection in the United States and Dependent Areas, HIV Surveillance Report, 2011. |
| [7] | [ B. Coburn,S. Blower, A major HIV risk factor for young men who have sex with men is sex with older partners, J. Acquir. Immune Defic. Syndr., 54 (2010): 113-114. |
| [8] | [ M. G. Cojocaru,L. B. Jonker, Existence of solutions to projected differential equations on Hilbert spaces, Proceedings of the American Mathematical Society, 132 (2004): 183-193. |
| [9] | [ Cojocaru, M. -G., Wild, E., On Describing the Solution Sets of Generalized Nash Games with Shared Constraints, submitted to: Optimization and Engineering, 2016. |
| [10] | [ M. G. Cojocaru,C. T. Bauch,M. D. Johnston, Dynamics of vaccination strategies via projected dynamical systems, Bulletin of mathematical biology, 69 (2007): 1453-1476. |
| [11] | [ M. G. Cojocaru, Dynamic equilibria of group vaccination strategies in a heterogeneous population, Journal of Global Optimization, 40 (2008): 51-63. |
| [12] | [ J. P. Dodds,A. Nardone,D. E. Mercey,A. M. Johnson, Increase in high risk sexual behaviour among homosexual men, London 1996-8: Cross sectional, questionnaire study, BMI, 320 (2000): 1510-1511. |
| [13] | [ F. Facchinei,A. Fischer,V. Piccialli, On generalized Nash games and variational inequalities, Operations Research Letters, 35 (2007): 159-164. |
| [14] | [ F. Fachinei,C. Kanzow, Generalized Nash Equilibrium Problems, Ann Oper Res, 175 (2010): 177-211. |
| [15] | [ J. W. Friedman, Game theory with applications to economics, Oxford University Press New York, 1990. |
| [16] | [ D. Gabay,H. Moulin, On the uniqueness and stability of Nash-equilibria in noncooperative games, Applied Stochastic Control in Econometrics and Management Science, 130 (1980): 271-293. |
| [17] | [ R. H. Gray,M. J. Wawer,R. Brookmeyer,N. K. Sewankambo,D. Serwadda,F. Wabwire-Mangen,T. Lutalo,X. Li,T. VanCott,T. C. Quinn, Probability of HIV-1 transmission per coital act in monogamous, heterosexual, HIV-1-discordant couples in Rakai, Uganda, The Lancet., 357 (2001): 1149-1153. |
| [18] | [ D. Greenhalgh,F. Lewis, Stochastic models for the spread of HIV amongst intravenous drug users, Stochastic Models, 17 (2001): 491-512. |
| [19] | [ T. B. Hallett,S. Gregson,O. Mugurungi,E. Gonese,G. P. Garnett, Assessing evidence for behaviour change affecting the course of HIV epidemics: a new mathematical modelling approach and application to data from Zimbabwe, Epidemics, 1 (2009): 108-117. |
| [20] | [ Y. H. Hsieh,C. H. Chen, Modelling the social dynamics of a sex industry: Its implications for spread of HIV/AIDS, Bulletin of Mathematical Biology, 66 (2004): 143-166. |
| [21] | [ J. Hofbauer and K. Sigmund, Evolutionary Games and Population Dynamics, Cambridge University Press, 1998. |
| [22] | [ D. M. Huebner,M. A. Gerend, The relation between beliefs about drug treatments for HIV and sexual risk behavior in gay and bisexual men, Annals of Behavioral Medicine, 4 (2001): 304-312. |
| [23] | [ T. Ichiishi, Game Theory for Economic Analysis, New York: Academic Press, New York, 1983. |
| [24] | [ D. Kinderlehrer,D. Stampacchia, null, An Introduction to Variational Inequalities and their Application, Academic Press, New York, 1980. |
| [25] | [ K. Nabetani, P. Tseng and M. Fukushima, Parametrized Variational Inequality Approaches to Generalized Nash Equilibrium Problems with Shared Constraints, (Technical Report 2008), Department of Applied Mathematics and Physics, Graduate School of Informatics, Kyoto University, Japan. |
| [26] | [ J. F. Nash, Equilibrium points in n-person games, CMS, 2 (2005): 21-56. |
| [27] | [ J.-S. Pang,M. Fukushima, Quasi-variational inequalities, generalized Nash equilibria, and multi-leader-follower games, CMS, 2 (2005): 21-56. |
| [28] | [ R. H. Remien,P. N. Halkitis,A. O'Leary,R. J. Wolitski,C. A. Gómez, Risk perception and sexual risk behaviors among HIV-positive men on antiretroviral therapy, AIDS and Behavior, 9 (2005): 167-176. |
| [29] | [ J. B. Rosen, Existence and uniqueness of equilibrium points for concave n-person games, Econometrica, 33 (1965): 520-534. |
| [30] | [ K. D. Schroeder,F. G. Rojas, A game theoretical analysis of sexually transmitted disease epidemics, Rationality and Society, 14 (2002): 353-383. |
| [31] | [ R. J. Smith,S. M. Blower, Could disease-modifying HIV vaccines cause population-level perversity?, The Lancet Infectious Diseases, 4 (2004): 636-639. |
| [32] | [ J. M. Stephenson,J. Imrie,M. M. D. Davis,C. Mercer,S. Black,A. J. Copas,G. J. Hart,O. R. Davidson,I. G. Williams, Is use of antiretroviral therapy among homosexual men associated with increased risk of transmission of HIV infection?, Sexually Transmitted Infections, 79 (2003): 7-10. |
| [33] | [ S. Tully,M. G. Cojocaru,C. T. Bauch, Coevolution of risk perception, sexual behaviour, and HIV transmission in an agent-based model, Journal of theoretical biology, 337 (2013): 125-132. |
| [34] | [ P. T. Harker, Generalized Nash games and quasi-variational inequalities, European Journal of Operational Research, 54 (1991): 81-94. |
| [35] | [ S. Tully, M. G. Cojocaru and C. T. Bauch, Sexual behaviour, risk perception, and HIV transmission can respond to HIV antiviral drugs and vaccines through multiple pathways, Scientific Reports, 5(2015). |
| [36] | [ J. X. Velasco-Hernandez,Y. H. Hsieh, Modelling the effect of treatment and behavioral change in HIV transmission dynamics, Journal of mathematical biology, 32 (1994): 233-249. |
| [37] | [ J. Von Neumann,O. Morgenstern, Theory of games and economic behavior, Bulletin of American Mathematical Society, 51 (1945): 498-504. |
| [38] | [ U. S. Census Bureau, Current Population Survey: Annual Social and Economic Supplement, 2012. Available at: https://www.census.gov/topics/population.html, (Accessed: Aug 2013). |
| [39] | [ University of Western News, HIV vaccine produces no adverse effects in trials, 2013, Available at: http://communications.uwo.ca/western_news/stories/2013/\September/hiv_vaccine_produces_no_adverse_effects_in_trials.html, (Accessed: Sept 2014). |
| [40] | [ Global Health Observatory, World Health Organization, 2014, Available at: http://www.who.int/gho/hiv/en/, (Accessed: Sept 2014). |
| [41] | [ Index Mundi, Zimbabwe Age structure, 2007, Available at: http://www.indexmundi.com/zimbabwe/age_structure.html, (Accessed: Aug 2014). |
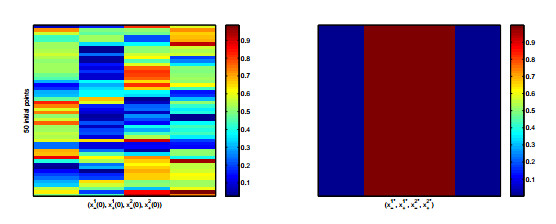
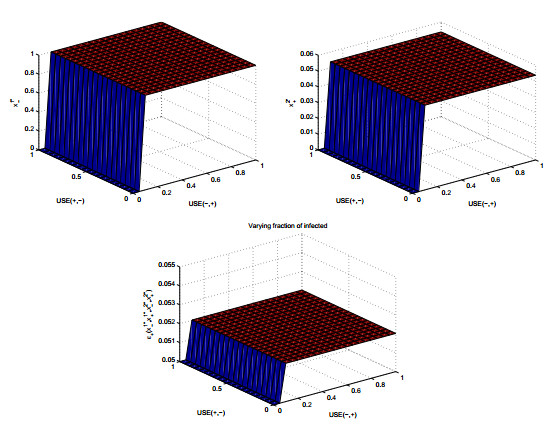
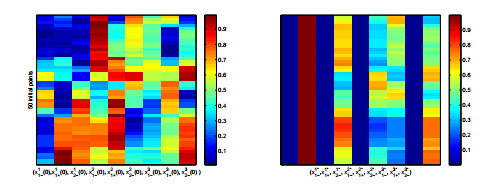
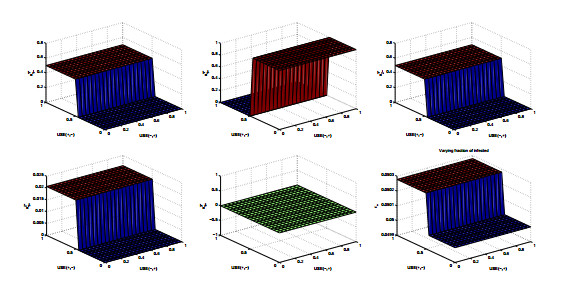
Figures(8) / Tables(2)

Stephen Tully, Monica-Gabriela Cojocaru, Chris T. Bauch. Multiplayer games and HIV transmission via casual encounters[J]. Mathematical Biosciences and Engineering, 2017, 14(2): 359-376. doi: 10.3934/mbe.2017023







 DownLoad:
DownLoad: