Advection and cross-diffusion terms are obtained as dispersal strategies of biological species. The focus of the paper is their connection to a given population dynamics. In particular, meaningful parameter regimes as biological dispersal are obtained. Eventually, we obtain a systematic approach to construct an advection or a cross-diffusion term from a given population dynamics and find meaningful parameter regimes as biological advection and cross-diffusion.
Citation: Jaywan Chung, Yong-Jung Kim, Ohsang Kwon, Chang-Wook Yoon. Biological advection and cross-diffusion with parameter regimes[J]. AIMS Mathematics, 2019, 4(6): 1721-1744. doi: 10.3934/math.2019.6.1721
Advection and cross-diffusion terms are obtained as dispersal strategies of biological species. The focus of the paper is their connection to a given population dynamics. In particular, meaningful parameter regimes as biological dispersal are obtained. Eventually, we obtain a systematic approach to construct an advection or a cross-diffusion term from a given population dynamics and find meaningful parameter regimes as biological advection and cross-diffusion.

| [1] | R. S. Cantrell and C. Cosner, Spatial ecology via reaction-diffusion equations, Wiley Series in Mathematical and Computational Biology, John Wiley & Sons Ltd., Chichester, 2003. |
| [2] | J. Cebrian, Energy flows in ecosystems, Science, 349 (2015), 1053-1054. |
| [3] | E. Cho and Y.-J. Kim, Starvation driven diffusion as a survival strategy of biological organisms, Bull. Math. Biol., 75 (2013), 845-870. |
| [4] | D. Cohen and S. A. Levin, Dispersal in patchy environments: the effects of temporal and spatial structure, Theoret. Population Biol., 39 (1991), 63-99. |
| [5] | L. Desvillettes, Y.-J. Kim, A. Trescases, et al. A logarithmic chemotaxis model featuring global existence and aggregation, Nonlinear Analysis: Real World Applications, 50 (2019), 562-582. |
| [6] | U. Dieckman, B. O'Hara and W. Weisser, The evolutionary ecology of dispersal, Trends Ecol. Evol., 14 (1999), 88-90. |
| [7] | J. Dockery, V. Hutson, K. Mischaikow, et al. The evolution of slow dispersal rates: a reaction diffusion model, J. Math. Biol., 37 (1998), 61-83. |
| [8] | I. A. Hatton, K. S. McCann, J. M. Fryxell, et al. The predator-prey power law: Biomass scaling across terrestrial and aquatic biomes, Science, 349 (2015). |
| [9] | R. Holt and M. McPeek, Chaotic population dynamics favors the evolution of dispersal, The American Naturalist, 148 (1996), 709-718. |
| [10] | V. Hutson, K. Mischaikow and P. Poláčik, The evolution of dispersal rates in a heterogeneous time-periodic environment, J. Math. Biol., 43 (2001), 501-533. |
| [11] | M. Johnson and M. Gaines, Evolution of dispersal: Theoretical models and empirical tests using birds and mammels, Ann. Rev. Ecol. Syst., 21 (1990), 449-480. |
| [12] | M. Keeling, Spatial models of interacting populations, advanced ecological theory: Principles and applications, J. McGlade, ed. Blackwell Science, Oxford, 1999. |
| [13] | E. F. Keller and L. A. Segel, Model for chemotaxis, J. Theor. Biol., 30 (1971), 225-234. |
| [14] | E. F. Keller, L. A. Segel, Traveling bands of chemotactic bacteria: A theoretical analysis, J. Theor. Biol., 30 (1971), 235-248. |
| [15] | Y.-J. Kim, O. Kwon and F. Li, Evolution of dispersal toward fitness, Bull. Math. Biol., 75 (2013), 2474-2498. |
| [16] | Y.-J. Kim, O. Kwon and F. Li, Global asymptotic stability and the ideal free distribution in a starvation driven diffusion, J. Math. Biol., 68 (2014), 1341-1370. |
| [17] | Y.-J. Kim and H. Seo, Model for heterogeneous diffusion, SIAM Appl. Math., 2019. |
| [18] | Y.-J. Kim, H. Seo and C. Yoon, Asymmetric dispersal and evolutional selection in two-patch system, Discrete Contin. Dyn. Syst., 2019. |
| [19] | K.-Y. Lam and W.-M. Ni, Limiting profiles of semilinear elliptic equations with large advection in population dynamics, Discrete Contin. Dyn. Syst., 28 (2010), 1051-1067. |
| [20] | M. McPeek and R. Holt, The evolution of dispersal in spatially and temporally varying environments, The American Naturalist, 140 (1992), 1010-1027. |
| [21] | J. D. Murray, Non-existence of wave solutions for the class of reaction-diffusion equations given by the Volterra interacting-population equations with diffusion, J. Theoret. Biol., 52 (1975), 459-469. |
| [22] | T. Nagylaki, Introduction to theoretical population genetics, Biomathematics, vol. 21, SpringerVerlag, Berlin, 1992. |
| [23] | W.-M. Ni, The mathematics of diffusion, CBMS-NSF Regional Conference Series in Applied Mathematics, vol. 82, Society for Industrial and Applied Mathematics (SIAM), Philadelphia, PA, 2011. |
| [24] | A. Okubo and S. A. Levin, Diffusion and ecological problems: modern perspectives, second ed., Interdisciplinary Applied Mathematics, vol. 14, Springer-Verlag, New York, 2001. |
| [25] | J. G. Skellam, Some phylosophical aspects of mathematical modelling in empirical science with special reference to ecology, Mathematical Models in Ecology, Blackwell Sci. Publ., London, 1972. |
| [26] | J. G. Skellam, The formulation and interpretation of mathematical models of diffusionary processes in population biology, The mathematical theory of the dynamics of biological populations, Academic Press, New York, 1973. |
| [27] | J. M. J. Travis and C. Dytham, Habitat persistence, habitat availability and the evolution of dispersal, Proc. Roy. Soc. London B., 266 (1999), 723-728. |
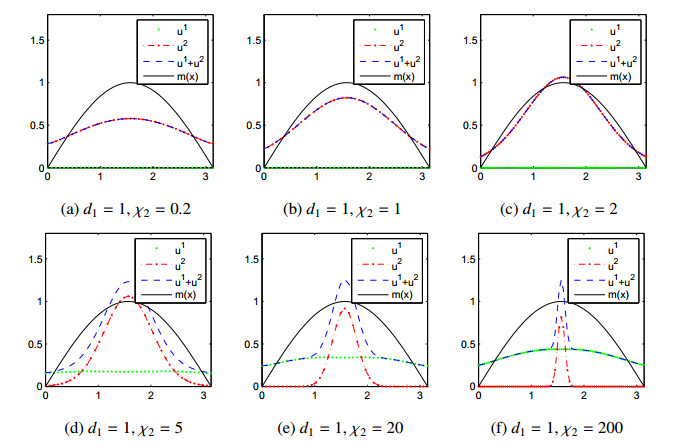
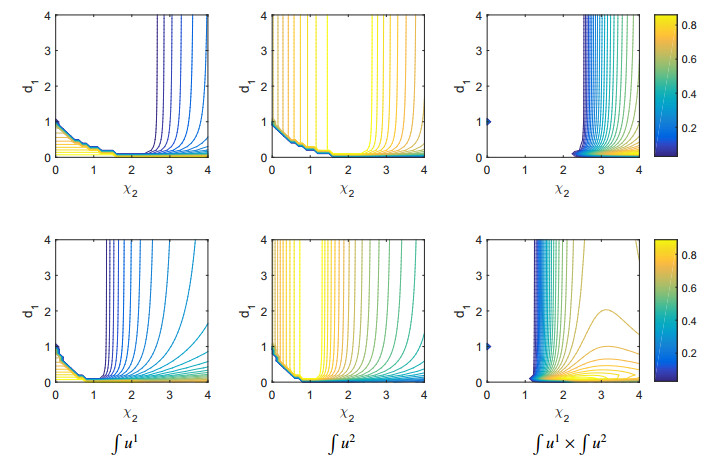
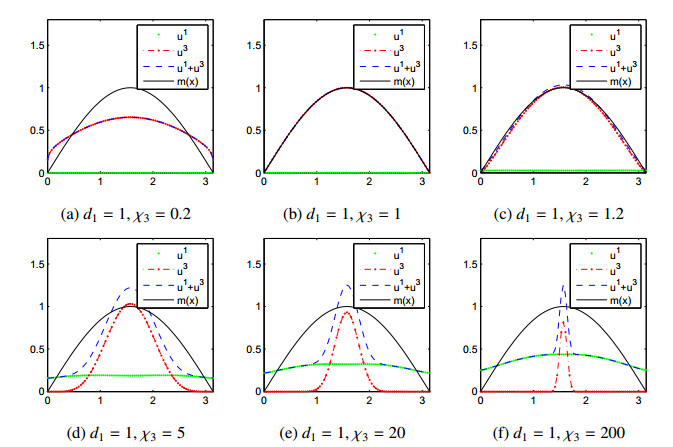
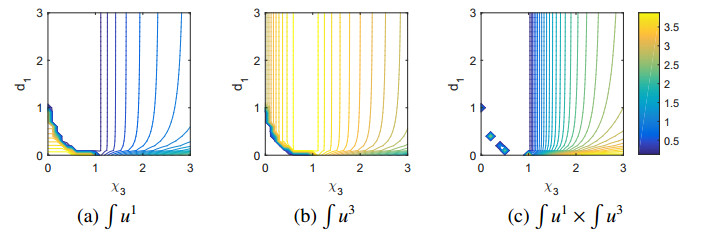
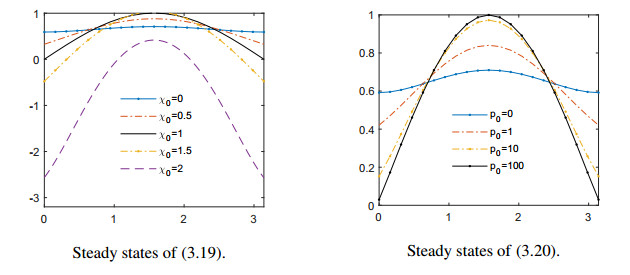
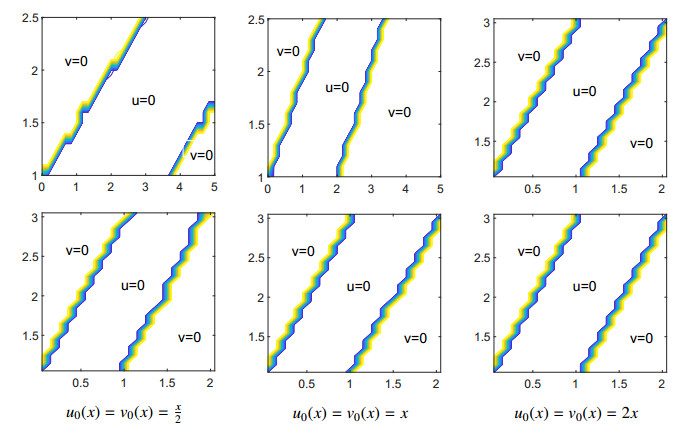
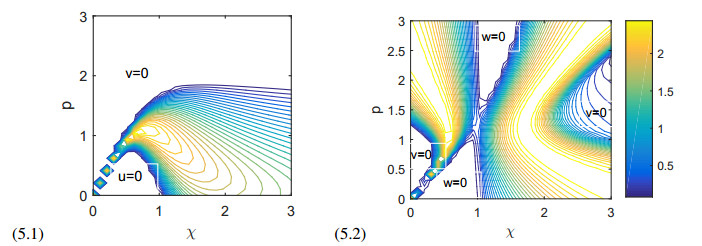
Figures(11)

Jaywan Chung, Yong-Jung Kim, Ohsang Kwon, Chang-Wook Yoon. Biological advection and cross-diffusion with parameter regimes[J]. AIMS Mathematics, 2019, 4(6): 1721-1744. doi: 10.3934/math.2019.6.1721







 DownLoad:
DownLoad: