Citation: Enrico Troiani. Analytical evaluation of the Stress Intensity Factor in stiffened sheets with multiple side damage[J]. AIMS Materials Science, 2016, 3(4): 1615-1622. doi: 10.3934/matersci.2016.4.1615

| [1] | Goranson U (2007) Damage tolerance facts and fiction. International conference on damage tolerance of aircraft structures, Delft University of Technology, Delft, the Netherlands. |
| [2] | Nesterenko B, Nesterenko G (2011) Analysis of requirements on fatigue and damage tolerance for civil transport airplanes. In: Komorowski J editor, Proceedings of the 26th Symposium of the international committee on aeronautical fatigue 39–59. |
| [3] | Bellinger NC, Komorowski JP (1997) Corrosion pillowing stresses in fuselage lap joints. AIAA J 35: 317–320. |
| [4] | Federal Aviation Administration (2011) Damage Tolerance and Fatigue Evaluation of Structure. Advisory Circular AC 25.571-1D. |
| [5] |
Wang W, Rans C, Alderliesten RC, et al. (2015) Predicting the influence of discretely notched layers on fatigue crack growth in fiber metal laminates. Eng Fract Mech 145: 1–14. doi: 10.1016/j.engfracmech.2015.06.062

|
| [6] | Wang L, Chow WT, Kawai H, et al. (1996) Prediction of widespread fatigue damage threshold. Proceeding of the FAA/NASA Symposium on Continued Airworthiness of Aircraft Structures, Atlanta (GA), USA. |
| [7] | Gruber ML, Wilkins KE, Worden RE (1996) Investigations of fuselage structure subject to widespread fatigue damage. In: Bigelow C editor. Proceedings from the FAA/NASA Symposium on the Continued Airworthiness of Aircraft Structures 439–460. |
| [8] |
Galatolo R, Nilsson KF (2001) An experimental and numerical analysis of residual strength of butt-joints panels with multiple site damage. Eng Fract Mech 68: 1437–1461. doi: 10.1016/S0013-7944(01)00027-3
|
| [9] | Schijve J (1994) Some calculation on the stress distribution in an infinite sheet with a single crack and with periodic collinear cracks. Report LR-770, TUDelft, the Netherlands. |
| [10] | Molinari G, Meneghin I, Melega M, et al. (2012) Parametric damage tolerance design of metallic aeronautical stiffened panels. Aeronaut J 1182: 815–831. |
| [11] |
Bloom JM, Sanders JL (1966) The effect of a riveted stringer on the stress of a cracked sheet. ASME J Appl Mech 33: 561–570. doi: 10.1115/1.3625122
|
| [12] |
Tada H (1970) Westergaard stress functions for several periodic crack problems. Eng Fract Mech 2: 177–180. doi: 10.1016/0013-7944(70)90023-8
|
| [13] | Poe CC (1971) Stress intensity factor for a cracked sheet with riveted and uniformly spaced stringers. NASA TR R-358, Langley Research Center, Hampton (VA), USA. |
| [14] | Westergaard HM (1939) Bearing pressures and cracks. J Appl Mech 6: 49–53. |
| [15] | Eftis J, Leibowitz H (1972) On the modified Westergaard equation for certain plane crack problems. Int J Fracture 8: 383–392. |
| [16] | Carthwrigth DJ, Rooke DP (1976) Compendium of Stress Intensity Factors, HMSO, London, UK. |
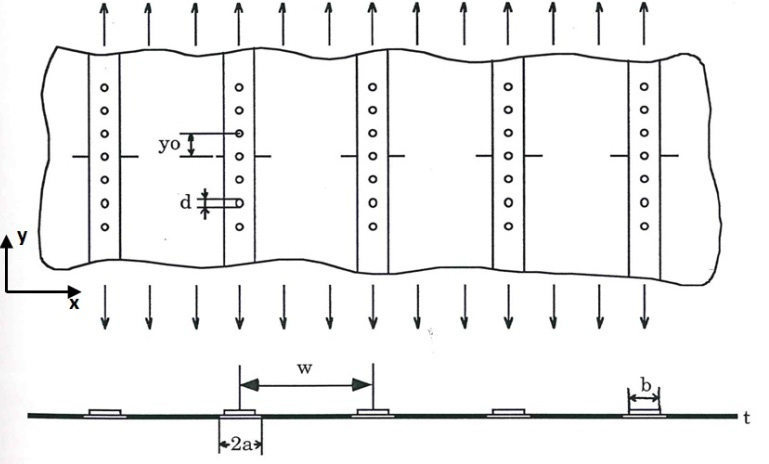
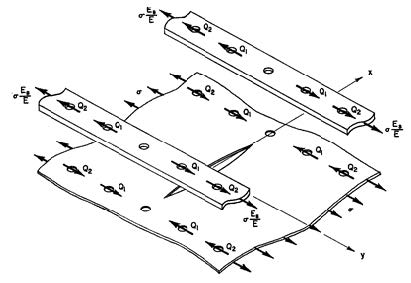
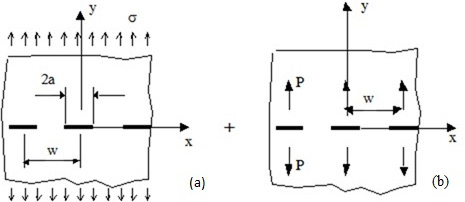
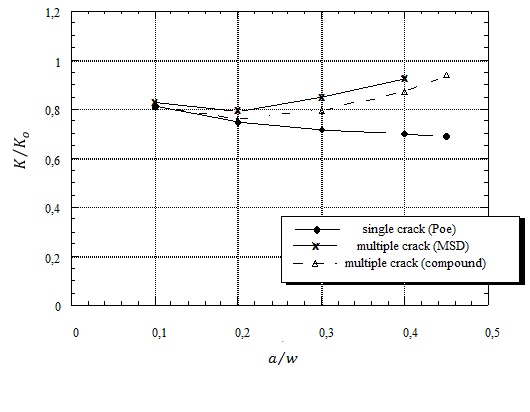
Figures(4)

Enrico Troiani. Analytical evaluation of the Stress Intensity Factor in stiffened sheets with multiple side damage[J]. AIMS Materials Science, 2016, 3(4): 1615-1622. doi: 10.3934/matersci.2016.4.1615







 DownLoad:
DownLoad: