




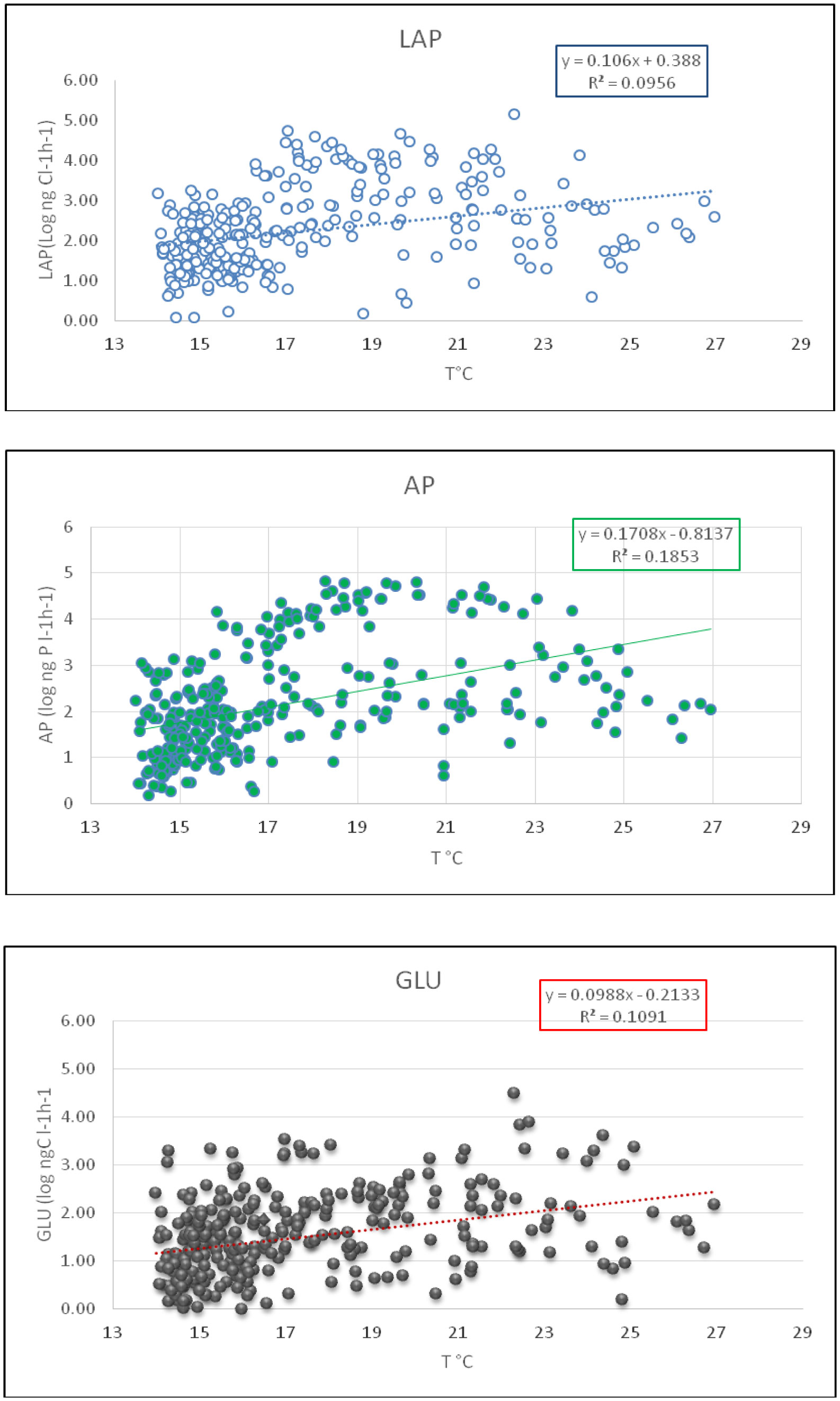
Figure 1.
Temperature in the euphotic layer of the Central Mediterranean versus LAP, AP and GLU activity rates from 1999 to 2013 (n = 320, P < 0.001).
Citation: Verónica Martínez-Fernández, Francisco Navarro. Rpb5, a subunit shared by eukaryotic RNA polymerases, cooperates with prefoldin-like Bud27/URI[J]. AIMS Genetics, 2018, 5(1): 63-74. doi: 10.3934/genet.2018.1.63
| [1] | Andrea Gärtner, Jutta Wiese, Johannes F. Imhoff . Diversity of Micromonospora strains from the deep Mediterranean Sea and their potential to produce bioactive compounds. AIMS Microbiology, 2016, 2(2): 205-221. doi: 10.3934/microbiol.2016.2.205 |
| [2] | Inês Barros, Hugo Froufe, George Marnellos, Conceição Egas, Jennifer Delaney, Michele Clamp, Ricardo Serrão Santos, Raul Bettencourt . Metatranscriptomics profile of the gill microbial community during Bathymodiolus azoricus aquarium acclimatization at atmospheric pressure. AIMS Microbiology, 2018, 4(2): 240-260. doi: 10.3934/microbiol.2018.2.240 |
| [3] | Nikolina-Alexandra Xaxiri, Eleni Nikouli, Panagiotis Berillis, Konstantinos Ar. Kormas . Bacterial biofilm development during experimental degradation of Melicertus kerathurus exoskeleton in seawater. AIMS Microbiology, 2018, 4(3): 397-412. doi: 10.3934/microbiol.2018.3.397 |
| [4] | Camilo Muñoz, Fernando G. Fermoso, Mariella Rivas, Juan M. Gonzalez . Hydrolytic enzyme activity enhanced by Barium supplementation. AIMS Microbiology, 2016, 2(4): 402-411. doi: 10.3934/microbiol.2016.4.402 |
| [5] | George P. Stamou, Sotiris Konstadinou, Nikolaos Monokrousos, Anna Mastrogianni, Michalis Orfanoudakis, Christos Hassiotis, Urania Menkissoglu-Spiroudi, Despoina Vokou, Efimia M. Papatheodorou . The effects of arbuscular mycorrhizal fungi and essential oil on soil microbial community and N-related enzymes during the fungal early colonization phase. AIMS Microbiology, 2017, 3(4): 938-959. doi: 10.3934/microbiol.2017.4.938 |
| [6] | O. Roger Anderson . Marine and estuarine natural microbial biofilms: ecological and biogeochemical dimensions. AIMS Microbiology, 2016, 2(3): 304-331. doi: 10.3934/microbiol.2016.3.304 |
| [7] | Mohammad Zahangeer Alam, Malancha Dey (Roy) . The reduction of abiotic stress in food crops through climate-smart mycorrhiza-enriched biofertilizer. AIMS Microbiology, 2024, 10(3): 674-693. doi: 10.3934/microbiol.2024031 |
| [8] | Ellie Sayers, Alex MacGregor, Simon R. Carding . Drug-microbiota interactions and treatment response: Relevance to rheumatoid arthritis. AIMS Microbiology, 2018, 4(4): 642-654. doi: 10.3934/microbiol.2018.4.642 |
| [9] | Amina Manni, Abdelkarim Filali-Maltouf . Diversity and bioprospecting for industrial hydrolytic enzymes of microbial communities isolated from deserted areas of south-east Morocco. AIMS Microbiology, 2022, 8(1): 5-25. doi: 10.3934/microbiol.2022002 |
| [10] | Denisse Galarza–Verkovitch, Onur Turak, Jutta Wiese, Tanja Rahn, Ute Hentschel, Erik Borchert . Bioprospecting for polyesterase activity relevant for PET degradation in marine Enterobacterales isolates. AIMS Microbiology, 2023, 9(3): 518-539. doi: 10.3934/microbiol.2023027 |
Marine microorganisms are the most abundant organisms in the oceans; in spite of their small size, they account for a significant fraction of living planktonic biomass and play a major role in biogeochemical processes in aquatic ecosystems [1]. In oligotrophic water bodies the role of microorganisms becomes even more important, because in such environments with low nutrient concentrations, prokaryotic heterotrophs compete with phytoplankton for the uptake of the low amounts of available nutrients, mostly derived by regeneration processes [2].
Heterotrophic bacteria contribute to nutrient cycles through the production of new bacterial biomass (bacterial carbon production) which acts as a temporary Carbon storage, influencing the amount of particulate organic matter (POM) available to higher trophic levels or exported to deeper waters. In fact, through the carbon uptake, prokaryotes convert the organic matter from dissolved to particulate phase, creating ‘food’ for large sized organisms within the ‘Microbial loop’ [3].
Large biomasses of photosynthetic microorganisms present in the oceans remove huge quantities of atmospheric carbon. Then, through the Microbial Carbon Pump mechanism [4],[5], microorganisms export carbon to the deep ocean, producing long-lived organic carbon. The ability of microorganisms to produce or degrade this carbon is expected to change according to the variations of environment in which the microbes are living.
Understanding how complex organic matter is transformed and remineralized by marine heterotrophs, and how this activity varies geospatially, is thus fundamental to our knowledge of the microbially-mediated carbon cycle on a global scale.
The organic matter present in marine waters has a heterogeneous matrix, composed of some labile materials such as proteins, simple sugars and fatty acids and other more refractory compounds, less prone to degradation. Indeed, heterotrophic bacteria can assume directly molecules with low molecular weight (below 600 daltons such as amino acids, simple sugars, fatty acids), sufficiently small to be transported across cell membranes, while organic polymers require to be first hydrolyzed into monomers or low- molecular weight compounds before their incorporation into the bacterial cell [6],[7]. The enzymatic hydrolysis of organic polymers by means of specific enzymes is generally considered as a key step in regulating organic matter utilization and its further processing [8].
Describing the variability in extracellular enzymatic activity in the marine environment, and studying the factors that drive this activity, is necessary to understand the global dynamics of the carbon cycle [2],[8],[9].
The Mediterranean Sea is generally recognized to be an oligotrophic basin with several distinctive features such as high summer evaporation and limited freshwater input, which make it particularly sensitive to climate changes [10]. Its main characteristics are:
Marine ecosystems play an important role in regulating climate, since they are a reservoir of 50% of anthropogenic carbon emissions, but at the same time, climate change affects natural systems. Understanding how microbial communities are affected by multiple climate change drivers is important to make accurate predictions of ecosystem evolution in response to changing climate scenarios [17].
Studies are currently performed to show that marine microbes could serve as sentinels to monitor climate change effects on marine ecosystems. Key ecosystem processes involved in Carbon cycle, like bacterial production, respiration, bacterial growth and losses by viral lysis and grazing, trophic interactions, all regulated by temperature, are likely to change in a warmer ocean, but the direction and the amplitude of phenomena are still a matter of debate [18]. This is particularly true for the Mediterranean Sea, reported to be one of the most oligotrophic ecosystems in the world [10].
Global warming is likely to reach 1.5 °C between 2030 and 2052 if it continues to increase at the current rate [19]. Many land and ocean ecosystems and some of the services they provide have already changed. The Mediterranean Sea levels will change and its circulation patterns will be modified [20].
Recent studies have reported a warming trend throughout the Mediterranean Sea, with temperature increases ranging from 0.017 °C yr−1 (in the mid-western Ionian sub-basin) to 0.05 °C yr−1 (north-east of the Levantine sub-basin). In the western Mediterranean Sea an increase of 0.032 °C yr−1 was reported; seasonal changes and spatial differences among its sub-basins were also observed [21].
The climate change occurring in the Mediterranean Sea may produce a number of evident direct and indirect effects on this marine ecosystem. The rapid heating of the water, especially since the '80s, is profoundly changing plankton biodiversity and food webs due to variations in the abundance and distribution of species [13]. Furthermore, as a direct effect, global warming may result in increased surface stratification, causing changes in thermohaline circulation and stimulating the rates of biological processes [18],[22]. However, predicting the effects on microbial abundance and metabolism of a warmer ocean is a current hot topic issue and the magnitude and implications of microbial response to climate changes remain still mostly unknown [17],[23],[24].
Sea temperature increase could influence the metabolism of heterotrophic microorganisms, causing cause changes in species distribution and in patterns of extracellular enzymatic activities that act both in surface and in deeper ocean layers.
Indeed, in the Mediterranean basins, increased enzymatic activities and decomposition rates have recently been observed [2012–2013] in the surface layer in coincidence with summer water warming [25],[26]. The Authors hypothesized that an important link between temperature and microbial enzymatic activities occurred in the surface layer of this area, with high amounts of organic matter utilized by prokaryotes mostly in warm periods, similarly to what found in other temperate regions by Hoppe et al. [27].
Other environmental factors that co-vary with temperature, such as nutrient concentration or primary productivity may themselves play a larger effect on bacterial activity [17],[18]. In fact, different environmental variables can influence each one of the microbial size components of food web and consequently the processes associated with global changes could be accelerated or compensated.
To date, no report on Mediterranean microbial community response to ocean acidification is available; in mesocosm experiments the effect of an increase in dissolved CO2 on the microbial communities in two opposite seasons was studied [28],[29].
Ocean acidification caused by the rising of CO2 produces changes in water pH; enzymes are sensitive to pH changes as they exhibit their activity in a specific pH range. However, the extracellular enzymes leucine aminopeptidase and glucosidase were not found to show significant changes when the pH decreased from 8.2 to 7.8 [17].
In order to better understand the factors driving hydrolysis patterns, this study investigated the relationship of enzyme activity rates with environmental variables, as well as their spatial and temporal variability. The link of heterotrophic enzyme activity with climatic changes led us to make predictions towards a major consumption and successive utilization of labile organic carbon derived from autochthonous (primary producers) or allochthons sources, so indicating that a heterotrophic metabolism could prevail in future.
The determination of enzymatic activities is fundamental to quantify microbial contribution to the turnover of organic matter [2]. The process of microbial decomposition is generally considered as a bottleneck in organic matter utilization, because the activities of microbial enzymes determine which substrates are hydrolysed and transported into the cells for further processes or respired and which substrates remain untouched and are transported to other areas or buried into sediments [6],[8].
For the field measurement of enzymatic activities, the fluorimetric method is a widespread technique generally used in aquatic environments, even if specific knowledge about the macromolecular structure of the organic matter in the ocean is lacking. According to the Hoppe's method [30], specific artificial substrates are widely used to measure potential enzyme activities in seawater and sediments. These substrate analogues consist of a monomer [an amino acid or a monosaccharide analogue] linked to a fluorophore, such as L-leucine-7-amido-4-methylcoumarin hydrochloride (leucine-MCA), used to measure leucine aminopeptidase; 4-methylumbelliferyl-β-D-glucopyranoside (MUF-β-glucose), and methylumbelliferyl-phosphate, used to measure β-glucosidase and phosphatase activities respectively. Many other fluorogenic substrates can be used to study chitinase, lipase, α-glucosidase, endopeptidase, ecc. [6].
When the link between the monomer and the fluorophore is cleaved by the specific enzyme, a fluorescent signal is emitted in coincidence with the release of the fluorophore, so that the hydrolysis of the fluorogenic substrate is measured by an increase in fluorescence over time.
Using the multiple substrate concentration procedure, measurements of the velocity of hydrolysis recorded at increasing substrate concentrations allow to calculate the maximum velocity of the reaction (Vmax), that is a measure of the potential enzyme activity of the microbial community present in the sample.
According to the kinetic approach, it is possible to determine also the Michaelis-Menten constant (Km), which expresses the affinity of an enzyme for its substrate [6],[30].
The fluorogenic assay is sensitive and relatively rapid, and the substrates analogues are usually used for comparisons among sites and studies. However, their concentrations routinely used in laboratory hardly reproduce the in situ conditions, because in situ the substrates are more diversified and their concentrations are generally unknown. A further limitation is that the enzyme assay does not discriminate among the different types of each extracellular enzyme potentially present in the environmental sample and measures only the activities of exohydrolase enzymes which cleave the terminal end of oligomers and not the endo-acting enzymes [2]. Based on these considerations, some recent studies [31] have suggested a conversion of ‘potential’ enzymatic activity into ‘actual’ rates of in situ hydrolysis of the assayed organic polymer.
Microorganisms are able to respond to changes in nutrient and substrate availability in the ecosystems by modifying their enzymatic profiles directly or through changes of microbial community composition; due to the plasticity of microbial metabolism, the determination of enzyme activity rates is considered a good indicator of environmental trophic conditions [9],[17].
During a study of the bacterial enzymatic potential in the north Adriatic Sea in different seasonal conditions, changes of bacterial community were observed. The main enzymatic activities of the bacterial community were peptidases, lipases and phosphatases, with a percentage increase in the hydrolytic potential during summer. Polysaccharides were scarcely hydrolysed by the bacterial community (average 20%) with a different seasonal cycle (alfa-glucosidase was more represented in the cold months, rather than in the warm months). These observations were attributable to succession of bacterial genera that possessed different enzymatic spectra. In fact, many bacterial groups possess peptidase, lipase and phosphatase, while chitinolytic enzymes and glucosidases are present in a limited number of genera only [32]. Other studies with molecular techniques evidenced changes in bacterial assemblage structure between summer and winter-spring periods [33]. However, the organic substances do not have all the same rate of degradation: some, depending on their biochemical composition and molecular structure, are demolished faster, while others require a long time. The lipids are generally the most easily assimilable organic component. The proteins and their derivatives are easily degradable. Carbohydrates make up a large part of the food reserve [8].
Extracellular enzymes can be found associated to cells or dissolved in the surrounding water. The free enzymes, defined as enzymes whose activity can be detected after filtration through a 0.2 µm pore-sized membrane, are recently studied in several environments. In many marine environments cell-free enzymes constitute more than half of the total activity [34]–[37]. They derive from active cells, as a consequence of viral lysis or protist grazing [38]. The importance of free enzymes for carbon cycling depends in part on the temporal and spatial scales over which such enzymes retain activity. These time- and distance scales suggest that these enzymes can affect carbon cycling at sites quite far from their production site [8],[34]. The temperature increase affects also the fraction of free enzymes, reducing the ratio of dissolved to total enzyme activity [36]. Only a few number of investigations, however, have been carried out in the Mediterranean Sea to estimate the contribution of dissolved enzymes to total enzyme activities [36],[37],[40],[41].
The most widespread studies on microbial enzymes in seawater regard the measure of the activities of leucine aminopeptidase -LAP; beta-glucosidase -GLU and alkaline phosphatase -AP.
Protein containing particles are very abundant in sea water and constitute a C and N source for bacteria [2],[8],[41].
LAP is an exopeptidase, because it cuts the extreme aminoacid of a polypeptides chain. The total LAP activity is usually used as an indicator of protein hydrolysis, because LAP provides amino acids for cellular metabolism and increases the availability of N; however, the interpretation of LAP activity is complex because this enzyme plays a dual role, being important in both carbon and nitrogen cycles. LAP is an enzyme very widespread in many marine and brackish environments [42]–[45] and may be synthesized by bacteria and cyanobacteria, ciliates and zooplankton [6].
GLU activity is generally used as indicator of polysaccharide hydrolysis. It is specific for the hydrolysis of cellobiose, a component of polymers such as cellulose and mucopolysaccharides; it contributes significantly to the recycling of both particulate and dissolved organic matter. Differently to LAP, it contributes to Carbon cycle, being able to hydrolyse more refractory organic compounds like polysaccharides and vegetal debris, derived from autochthonous or allochthons production.
Glucosidases are known to be ubiquitously present in seawater [6],[37]. In temperate zones GLU is usually lower than LAP; this can be explained by the presence of constitutive aminopeptidase during summer, when polysaccharides and protein degradation are not correlated [40],[46].
Beside beta-glucosidase, which hydrolyses the beta -D- link of cellobiose, alfa -glucosidase (cleaving alfa linked D-glucose) and more different enzymes are active in polysaccharides hydrolysis, as reported in many studies [7],[45].
Phosphatases are also enzymes widely spread in aquatic environments. AP is involved in the hydrolysis of dissolved organic phosphates (DOP), with regeneration of inorganic phosphate [47]. In the epipelagic layer, the amount of total AP of bacterial origin could also be over-estimated since this enzyme is produced by both prokaryotes (bacteria and cyanobacteria) and eukaryotes (phyto- and zoo- plankton) [39]. AP is usually considered an inducible enzyme, being synthesized under low levels of PO4 and repressed when P becomes available. In fact, the expression of ectoenzymes is regulated by a negative feedback, so the availability of inorganic P could have an inhibitory effect on AP synthesis [5]. Therefore, AP has been suggested as an index of P deficiency, because it is produced in great amounts when inorganic P becomes limited [48],[49]. It assumes great importance in very P- limited areas such as the Eastern Mediterranean Sea [49].
To this regard, in the very oligotrophic Levantine Sea a limiting factor for bacterial production was the availability of inorganic nutrients, especially phosphorus. How P limitation could affect microbial food web and influence carbon flow was investigated by Thingstad et al. [50]. However, while phosphorus is usually the limiting nutrient, nitrogen and carbon limitation or co-limitation also occur, and the type of limitation can vary with depth [49],[51]–[53]. Moreover, different mechanisms are involved in the N and P regeneration: AP activity is responsible for the direct mineralization of organic phosphate esters into orthophosphate, while LAP acts in the first steps of protein decomposition, with the release of dipeptides which follow further decomposition pathways before their final mineralization.
The presence of AP below the surface layer and at deep layers assumes a different meaning due to abundance in deep layers of mineralized nutrients such as P. It has been hypothesized that AP is not only regulated by inorganic P concentration, but it could be produced in C-limited conditions, contributing to both C and P cycles [47],[51].
The majority of the investigations of enzymatic activities in marine systems have been carried out in the water column. In marine environments, activities are typically higher in surface waters than at depth and vary between nearshore and offshore sites [26],[27].
Christian and Karl [54] suggested that patterns of peptide hydrolysis may differ between temperate and high latitude sites, increasing at high latitude, while carbohydrate hydrolysis at lower latitudes, similarly to that documented for polysaccharides [45].
The distribution of enzymatic activities in diverse Mediterranean sites, classified into coastal, offshore and frontal systems, is reported in Table 1. Differences in enzyme patterns were observed in relation to time (seasons and years), or space, between coastal and pelagic waters [46],[49]. Consequently, patterns of polymer hydrolysis rates change horizontally and vertically, among different water masses, along estuarine gradients [54]–[57].
Only few studies have considered the distribution of hydrolytic enzymes along longitudinal transects in the Mediterranean Sea: Van Wambeke et al. [51] reported LAP ranging from 1.1 to 9.3 nmol l−1h−1 without a clear East-West gradient; La Ferla et al. [56] and Zaccone et al [49] along a W-E longitudinal transect (Transmed cruise), found regional differences within a variety of Mediterranean provinces. Also Luna et al 2012 [59] showed significant differences between the West and Central East Mediterranean basins; the variance in LAP activity rates accounted for most of the observed total variance in enzyme distribution.
Aminopeptidase generally showed a decreasing gradient from more eutrophic coastal areas to oligotrophic waters [60]. In the East Mediterranean the range of variation of LAP activity (0.3–4.91 nmol l−1h−1 [49]) was similar to that reported in the NW Mediterranean [50] and in the Ionian Sea [46],[55], but lower than that observed in the Tyrrhenian Sea [62],[63]. Placenti et al. [64] in an Ionian pelagic area observed high LAP values (Table 1), similar to those recorded in coastal zones [65],[66]. Also in the Atlantic Ocean LAP values varied in a wide range [27],[34].
Subsurface peaks of LAP roughly corresponded to chlorophyll peaks and high rates of activities were observed in the Deep Chlorophyll Maximum (DCM), indicating the contribution of planktonic autotrophic organisms (cyanobacteria and phytoplankton) to the synthesis of this enzyme. The pico- and nano-sized organisms were the dominant fraction in the DCM, with a peculiar microbial food web. The water column stratification during summer resulted in an accumulation of particulate matter, with consequent concentration of prokaryotic biomass and metabolic activity [14],[40].
Generally low hydrolysis rates of GLU were observed in the Central Mediterranean Sea (Table 1), except in autumn along all the water column [63]. Differences in polysaccharide hydrolysis between surface and subsurface waters were observed by Arnostì [45] in other areas, suggesting substantial changes in the spectrum of substrates utilized by microorganisms.
Differences in AP activity levels among the water masses present in the water column were also observed [55]. In the Ionian Sea the decreasing trend of AP versus PO4 from the surface to deep waters was described by an exponential curve showing a significant correlation (R2 = 0.77, p < 0.05) and indicating that the abundance of inorganic P could have an inhibitory effect on AP activity. In the same area an inverse relationship with inorganic P was found [46] except in autumn [64]. Similarly, in the south Adriatic Sea an exponential curve described the vertical decreasing trend of AP versus PO4 [57]. In the north Adriatic Sea surface AP was significantly higher than at the bottom [37],[38].
In the Ionian Sea the maximum hydrolysis rates of LAP, GLU and AP were measured in the surface layer. Significant differences (ANOVA P > 0.05) among epipelagic layer, meso-, and bathypelagic waters were reported in this basin [46]. Depth profiles of aminopeptidase exhibited surface values 8 times higher than at depth. Several Authors observed this decreasing trend along the water column in relation to availability of organic carbon [49],[55],[56],[64],[67]. Also Luna et al. 2012 [59] showed significant differences among epi- meso- and bathypelagic layers (P < 0.001). Occasionally, an increasing trend of LAP towards deep layers was observed [63].
Heterotrophic microbes at depth are able of degrading organic matter (semi-labile DOC) that resists to the biological removal occurring in the surface ocean [45]. Additional deep sea measurements are from Misic and Fabiano [68] and Celussi et al [69]. A strong relationship linking enzyme activity rates with the presence of different water masses is a common finding of these studies [46],[49],[55],[57].
Depth profiles of bacterial abundance and metabolic rates extending to the deeper meso- and bathypelagic zones indicate that cell numbers decreased by a larger factor than enzyme activities, so that cell-specific enzyme activities were calculated to really compare with measurements taken in surface the observations obtained from deeper waters. These studies also found that enzyme activities calculated on a per-cell basis were generally higher at depth compared to surface ocean waters; this approach did not take into account the amount of active or non-viable cells, nevertheless it allowed to compare different environments [34].
The vertical distribution of enzyme activities normalized per cell showed a trend different for the three enzymes: LAP prevailed in the epi- and bathypelagic layers, whereas the importance of GLU increased at the mesopelagic layer [46]. In the deep zone of the Ionian Sea also Placenti et al [64] observed an increasing trend in the specific activity of deep prokaryotes; the importance of specific GLU activity, compared with LAP, indicated a preferential hydrolysis of more refractory material, such as mucopolysaccharides, compared to the protein fraction, which was preferentially hydrolysed in the surface layers [46],[63]. An increase in the specific enzyme activity with depth was also observed by Baltar et al. [67] who explained this result as an adaptation of enzyme activity to exploit the scarce amount of available organic matter or to its refractory nature.
The sea temperature increase in the Mediterranean Sea may have effects on heterotrophic microorganisms, and could reinforce the major role in organic matter decomposition or production, alternatively, in the carbon cycle [25]. According to Wohlers et al. [22], the temperature increase detected in surface waters affected the hydrological structure of water column, favouring water stratification and stimulating the biological processes. Weather dependent alterations of seawater temperature with implications on bacterial metabolism were also observed [26],[70]. As most of the extracellular enzymes are adaptive, environmental and biological factors such as temperature, pH and salinity, quality and quantity of organic matter may affect their activity rates. Physical and chemical factors partially explain the changes in potential enzymatic activities. This does not mean that necessarily there is a direct causal effect, but multiple factors concur to stimulate bacterial activity. The correlative approach led us to hypothesize that the increase of microbial processes on organic matter was linked to water warming. The meaning of this finding could be that the rising temperatures, as expected by water warming, may cause an increase in remineralisation–at the cost of photosynthetic production – with consequent predominance of a heterotrophic regime [26]. In fact, increasing temperature stimulates microbial growth, but cells require also substrate and nutrients, supplied by microbial activities.
The surface layer was particularly sensitive to temperature changes and seasons. In Figure 1 the enzymatic activities were plotted in relation to water temperatures in the epipelagic layer of Ionian Sea. The relationship was significant for all tested enzymes, with different slopes. In the meso- and bathy-pelagic layer the relationship with temperature was not significant, because the range of T varied from 13.30 to 15.00 °C, over the different seasons.
Moreover, the warming effects on microbial components have been explored in the Sicily Channel in relation to fishery resources. The results confirmed that a co-variation of prokaryotic activities and eggs distribution with temperature exists in summer, suggesting a reciprocal relationship among them [14],[40].
Different behaviour in prokaryotic abundance and microbial enzymatic activities between cold and warm periods was previously observed [46],[71]. Conversely, during winter the effect of T was scarce, because the epipelagic layer was mixed (ΔT = 0.13 °C). In this season the recycling of P by microbial activity assumed more importance in controlling prokaryotic biomass and activities [40]; this was in agreement to Cea et al. [71] that observed P limitation in the NW Mediterranean Sea.
In coastal and estuarine environments, the abiotic factors that influence microbiological processes may be the same, but change in their relative importance depending on the type of environment. However, particular elements can be recycled more quickly than others, causing decoupling in biogeochemical cycles. In these ecosystems the effect of T increase was higher than in pelagic areas, indicating a higher degree of correlation between bacterial activity and temperature. There was also a narrow co-variation with bacterial production and chlorophyll-a content [27],[42].
In the superficial waters of the Adriatic Sea a doubling of the peptidase activity has been observed in summer in comparison to the spring [70],[72]. Coastal ecosystems - which are subject to rapid changes of organic matter due to main current, riverine input, or eutrophication - support a wide spectrum and high rates of enzymatic activity [32],[43],[74]. In fact, in these environments micro-organisms respond rapidly to changes in nutrient and substrate availability modulating their enzymatic profiles in relation to new organic polymers and, in turn, changes in the specific composition of the microbial community affect the decomposition process [45].
The distribution of enzymatic activity and in particular of proteases in different marine and lacustrine environments highlighted a great activity in the superficial layers. An increase in enzyme activity may also occur near the bottom in relation to the available organic matter. In brackish waters, changes in prokaryotic metabolism did not depend on variations in the prokaryotic abundance, but rather on the stimulating effect played by seasonal trophic variations. In these small ecosystems seasonal changes in the local environmental parameters (trophic supply, CHL, POC, allochthons input, storm events) are more evident and significant seasonal variations in the patterns of microbial activity in relation to temperature, dissolved oxygen and trophic changes were found [45],[74]. The measured rates of enzymatic activities were consistently higher than that observed in coastal marine areas; moreover, positive correlations between temperature and LAP, AP, particulate organic matter were observed in Ganzirri Lake [45],[75].
Salinity is a significant factor affecting negatively the enzyme distribution in most areas of the Mediterranean Sea [49]. The effect of Salinity is due to other changes (like inorganic nutrient input, land derived organic load, primary production stimulation etc.) that the river water causes in coastal waters. [66],[70],[72].
A negative correlation among S and abundance and microbial activities was reported in several aquatic environments [17],[31],[43],[58],[72]. In fact, in the Adriatic Sea, where inputs of the Po river affect a wide coastal zone, the range of salinity was very different from that of pelagic areas. The LAP activity was concentrated in the surface layer where the stratification of the fluvial waters brings high amounts of organic matter, and decreased along a gradient of salinity from the coastal stations towards the open sea [73]. The activity values were 1.5–1.7 times greater in summer than in winter [71], with a trend that followed the trophic gradient towards the Po delta.
On the contrary, in the pelagic area of Sicily Channel salinity did not appear to be one of the most important physical-chemical parameters driving microbial metabolism and abundance. The salinity range measured in the Ionian Sea by Zaccone et al. [40] was very small (ΔS = 0.72 in summer and 0.76 in winter), like that observed by Monticelli et al. [58] in a coastal zone affected by a thermohaline front (ΔS = 0.81, ΔT = 3.44 °C).
In brackish environments a gradient of salinity and progressive segregation from the sea was observed in the coastal lagoon of Oliveri-Tindari, a very suitable study environment with ecological characteristics differing from each other (Marinello ponds) [60]. The marked spatial and temporal diversification of the ponds in relation to their trophic features (ranging from oligo- to eutrophic) was reflected in a various and unpredictable range of enzyme patterns. With respect to the ponds nearest to the sea, higher level of microbial activity (ranges: 0.55 to 4359.03 nM h−1, 0.15 to 76.44 nM h−1, 1.29 to 1548.06 nM h−1 for LAP, GLU and AP respectively) and a greater variability, due to seasonal course and terrestrial input, characterised the most ‘confined’ ponds. Lower enzyme activities were observed in less eutrophicated ponds. With regard to their seasonal distribution, all the enzymes examined showed a sharp increase in all ponds from March to June (spring peak), and a second peak in August–September [60].
In the Cape Peloro lagoon AP was 10.83 folds higher in summer-autumn than in winter-spring, while GLU in summer-autumn was 19.6 folds higher than in winter-spring. Moreover, during summer-autumn periods a prevalence of AP over the LAP and GLU activities was observed, indicating the high demand of P, that was quickly regenerated. On the contrary during winter-spring months the prevalence of LAP over other enzymes confirmed changes in prokaryotic metabolism [75].
The relative importance of LAP, GLU and AP reflects changes in the relative amounts of proteins, polysaccharides and organic phosphates, respectively. In surface waters, the magnitude of enzymatic activities is generally in the order aminopeptidase > phosphatase > glucosidase. A more rapid hydrolysis of leucine-MCA compared to the one of MUF-glucose indicates a more rapid remineralization of organic nitrogen than organic carbon during early diagenesis [2],[8]. This order of magnitude may change in relation to season, latitude (polar or temperate zones), quality of available organic matter in water column or sediments [17],[54]. Variation in the relative activities of different enzymes is suggested as a possible indicator of changes in the modalities of bacterioplankton nutrition [54].
The reciprocal ratios among enzyme activities in plankton communities have been focused by some Authors [76] who have suggested the ‘ecoenzymatic stoichiometry’ as a tool to connect the elemental stoichiometry of microbial biomass and organic matter to nutrient assimilation and microbial growth. According to this approach, the relative abundance of enzymes involved in C, N, and P cycles reflects the biogeochemical equilibrium between microbial biomass and the elemental composition of organic matter.
The reciprocal ratios between enzymatic activities (LAP/GLU and LAP/AP) reported by different Authors are shown in Table 1.
| Area | Depth (m) | Sites | season | T °C | S | LAP(nM h−1)* | GLU(nM h−1)* | AP(nM h−1)* | LAP/GLU molar ratio | LAP/AP molar ratio | References |
| NW Mediterranean | 10–2000 | DYFAMED area | spring | >12.70 | 38.40 | 1.78–0.23 | 1.12–0.22 | 1.6 | [61] | ||
| 10–2000 | DYFAMED area | fall | 1.44–0.45 | 1.5–1.2 | 0.37 | " | |||||
| NW Mediterranean | DYFAMED area | autumn 04 | 3.7–4.6 | 2.4–3.2 | [53] | ||||||
| Mediterranean | 5–200 | W-E transect | summer 99 | 22.7–24.9 | 37.28–39.01 | 1.1–9.3 | 0.62–1.65 | [51] | |||
| NW Mediterranean | 5 | Marseille Bay | annual 12-13 | 12.8–25.8 | 38.15 | 5.1–85.1 | 0.1–115.7 | 5.42 | [71] | ||
| Ligurian Sea | 5–200 | gulf of Genoa | summer 02 | 17–24 | >36.7 | 1.78–4.04 | 0.04–0.19 | 1.52 | 25-68 | [68] | |
| Ligurian Sea | 0.5 | Portofino | spring -summer 07 | 19.4 | 110.1–165.6 | 13.9–29.4 | [66] | ||||
| South Tyrrhenian Sea | 1–100 | Eolian Island | spring 04 | 1.9–2.5 | 0.04–0.08 | [65] | |||||
| Tyrrhenian sea | 25–3000 | winter 05 | 13–16 | 37.8–38.25 | 0.3–21.7 | 0.5–9.9 | [62] | ||||
| South Tyrrhenian Sea | 5–100 | summer 05 | 17.55 | 0.59 | 3.60 | 29.75 | 4.88 | [63] | |||
| " | 200–1000 | " | 23.71 | 0.80 | 0.78 | 29.64 | 30.40 | " | |||
| " | >1000 | " | 37.72 | 0.40 | 0.93 | 94.3 | 40.56 | " | |||
| Tyrrhenian sea | 10–200 | st. VECTOR | summer 07 | 16.79 | 38.08 | 0.31 | 0.31 | 1.16 | 1 | 0.27 | [49] |
| " | 500–1000 | " | " | 13.81 | 38.69 | 0.37 | 0.37 | 0.19 | 1 | 1.95 | " |
| " | 1500–3500 | " | " | 13.49 | 38.53 | 0.61 | 0.47 | 0.20 | 1.29 | 3.05 | " |
| Ionian Sea | 5–200 | pelagic | winter 98 | 15.17 | 38.59 | 1.92 | 0.19 | 1.30 | 9.91 | 0.67 | [55] |
| 400–1000 | " | 13.98 | 38.80 | 1.59 | 0.16 | 0.16 | 9.94 | 0.10 | " | ||
| 1100-4000 | pelagic | " | 13.92 | 38.76 | 0.51 | 0.06 | 0.92 | 8.81 | 1.79 | " | |
| Ionian Sea | 5–200 | pelagic | winter 99 | 15.79 | 38.65 | 5.40 | 4.85 | 9.97 | 1.12 | 0.54 | [26], [46] |
| 400–1000 | " | 13.60 | 38.73 | 2.83 | 0,70 | 1.90 | 4.05 | 1.49 | " | ||
| 2000–3300 | " | 13.39 | 38.71 | 5.97 | 0.49 | 11.97 | 12.17 | 0.50 | " | ||
| Ionian Sea | 5–200 | pelagic | autumn 00 | 17.47 | 38.68 | 1,48 | 1,79 | 99.86 | 0.83 | 0.02 | [26],[46] |
| 400–1000 | " | 13.77 | 38,76 | 1,58 | 0,51 | 52.27 | 3.11 | 0.30 | " | ||
| 2000–3300 | " | 13.79 | 38.73 | 1,53 | 2,86 | 35.41 | 0.53 | 0.04 | " | ||
| Ionian Sea | 5–200 | pelagic | summer 01 | 17.38 | 38.62 | 2,02 | 0,20 | 5,08 | 10.12 | 0.40 | [26],[46] |
| 400–1000 | " | 13.81 | 38.76 | 10,64 | 0,35 | 4,82 | 29.97 | 2.21 | " | ||
| 2000–3300 | " | 13.79 | 38.73 | 1,08 | 0,34 | 3,12 | 3.17 | 0.35 | " | ||
| Ionian Sea | 5–200 | pelagic area | autumn 04 | 17.14 | 38.44 | 4,95 | 2,04 | 4,06 | 2,43 | 1,22 | [26],[46] |
| 500–1000 | " | 13.96 | 38.80 | 3,60 | 0,34 | 1,63 | 10.46 | 2,21 | " | ||
| 1100– 4000 | " | 13.85 | 38.80 | 4,53 | 0,80 | 2.01 | 5.67 | 2,25 | " | ||
| Ionian Sea | 5–200 | pelagic area | spring 02 | 15.49 | 38.58 | 2,96 | 1,44 | 5.29 | 2.05 | 0.56 | [26],[46] |
| 500–1000 | " | 14.02 | 38.79 | 0,50 | 0,10 | 4.87 | 0,97 | " | |||
| 1100– 4000 | " | 13.84 | 38.74 | 0,33 | 0,09 | 0,16 | 3.70 | 2,03 | " | ||
| Ionian Sea | 0–150 | pelagic area | autumn 11 | >15.50 | <38.80 | 21.10 | 18.91 | 144.99 | 1.12 | 0.15 | [64] |
| 200-600 | " | " | < 15.00 | >39.00 | 20.84 | 15.03 | 14.26 | 1.39 | 1.46 | " | |
| >3000 | " | " | 13.42 | >38.72 | 20.39 | 12.94 | 12.21 | 1.58 | 1.67 | " | |
| Ionian Sea | 5–100 | pelagic area | summer 12 | 18.16 | 38.29 | 4.91 | 1.37 | 2.04 | 3,59 | 2,40 | [26],[40] |
| Ionian Sea | 5–100 | " | winter 13 | 14.97 | 38.45 | 1.49 | 0.27 | 16.74 | 5.53 | 0.09 | " |
| Ionian Sea | 1–60 | coastal station | autumn 01 | 19.26 | 38.53 | 207.29 | 11.28 | 717.8 | 18.37 | 0,29 | [79] |
| Atlantic ocean | 5–100 | Atlantic station | summer 07 | 0.38 | 0.27 | 0.22 | 1.41 | 1.73 | [49] | ||
| 200–1000 | " | 0.74 | 0.13 | 0.05 | 5.69 | 14.8 | " | ||||
| 1500–3000 | " | 0.57 | 0.25 | 0.01 | 2.28 | 57 | " | ||||
| West Mediterranean | 5–200 | pelagic area | summer 07 | 16.21 | 37.8 | 0.35 | 0.20 | 1.21 | 1.75 | 0.29 | [49] |
| 500–1000 | " | 13.36 | 38.57 | 0.21 | 0.32 | 0.19 | 0.66 | 1,11 | " | ||
| West Mediterranean | 1500–3500 | " | 13.29 | 38.50 | 0.36 | 0.25 | 0.20 | 1.44 | 1.8 | " | |
| East Mediterranean | 5–200 | summer 07 | 18.41 | 38.79 | 1.53 | 0.50 | 4.56 | 3.06 | 0.33 | [49] | |
| 500–1000 | " | 14.01 | 38.81 | 0.66 | 0.52 | 0.33 | 1.27 | 2.01 | " | ||
| 1500– 2900 | " | 13.95 | 38.75 | 1.22 | 0.58 | 0.78 | 2.10 | 1.56 | " | ||
| West Mediterranean | 5–200 | 2.18 | 0.88 | 0.51 | [59] | ||||||
| 500–1000 | 1.04 | 0.51 | 0.35 | " | |||||||
| West Mediterranean | 1500–3500 | 0.66 | 0.41 | 0.31 | " | ||||||
| East Mediterranean | 5–200 | 0.08 | 0.31 | 0.20 | [59] | ||||||
| 500–1000 | 0.09 | 0.10 | 0.13 | " | |||||||
| 1500– 2900 | 0.05 | 0.13 | 0.03 | " | |||||||
| North Adriatic Sea | gen-60 | coastal station | seasonal study | 2.2–1701 | 0.3–114 | 0.8 -1770 | [70] | ||||
| North Adriatic Sea | 1–60 | coastal station | spring-summer | 115– 67.5 | 2.1–5.2 | 13.6 | 22.14 | 4.96 | [73] | ||
| North Adriatic Sea | autumn | 2.04–4.64 | [38] | ||||||||
| North Adriatic Sea | 1–10 | coastal station | spring-autumn | 70–4640 | [37] | ||||||
| South Adriatic Sea | 5–100 | convection site | seasonal study | 13.8–15.6 | 38.71–38.74 | 0.01–7.86 | 0.003–3.5 | 0.05–31.96 | [57] | ||
| 100–800 | " | 13.6–13.6 | 38.74–38.77 | 0.01–2.66 | 0.003–0.98 | 0.003–6.47 | " | ||||
| " | 800–1200 | " | 12.9–13.1 | 38.72–38.73 | 0.01–0.15 | 0.003–0.95 | 0.08–17.96 | " | |||
| South Adriatic Sea | 0–30 | Gulf of Manfredonia | spring 03 | 18.44 | 37.92 | 13.99 | 1.13 | 43.32 | 12.4 | 0.32 | [58] |
| 0–30 | offshore waters | " | 17.25 | 38.73 | 31.18 | 0.48 | 43,28 | 72.51 | 0.72 | " | |
| >30 | " | " | 14.11 | 38.70 | 19.45 | 0.26 | 23.49 | 75.97 | 0.83 | " | |
| North Atlantic | surface | 4.2–24.9 | [27] | ||||||||
| South Atlantic | surface | 11.5–151.4 | " | ||||||||
| Atlantic ocean | 100–4500 | 0.2–13.8 | 0.05–0.25 | 0.14–1.3 | [34] | ||||||
| subtropical Atlantic | 100–4500 | 0.6–9.2 | 0.02–0.07 | 0.04–3.7 | [67] |
 DownLoad:
CSV
DownLoad:
CSV
Particularly, the LAP/GLU ratio indicates the ratios between the two processes of proteolysis and glycolysis and depends on the functional features of each ecosystem. High LAP/GLU ratios provide information on the flux of organic matter preferentially through proteins, while low ratios indicate a preferential flux of organic matter through polysaccharides [44]. Together with the common quality indexes (C/N ratio), this ratio provides information on the potential lability of the organic matter [44],[66].
LAP/GLU ratios higher than 1 are generally observed in coastal and pelagic environments and suggest the presence of fresh organic matter such as those produced by phytoplankton [2],[48]. In the Mediterranean Sea, positive ratios have been used as an indicator for nitrogen-rich versus carbon-rich substrates [68] (Table 1).
In the Ionian Sea, the LAP/GLU as molar ratio ranged from 0.8 to 10.1 in epipelagic waters, with high values in summer, indicating that microbial community uses LAP to have access to protein substrates as both C and N source [46],[64]. Along a Western-Eastern Mediterranean transect Zaccone et al [49] found the prevalence of LAP over GLU along a West–East transect in epi- and bathy-pelagic layers (Table 1).
High LAP/GLU ratios in deep Ionian waters were found in winter 1998 and 1999 [46],[55]. A seasonal trend of this ratio was observed by Azzaro et al [57] in the south Adriatic Sea, with higher values in April and June probably due to a high amount of proteins in the photic layer during productive phase. LAP/GLU ratio decreased during phytoplankton decay and the protein components of organic matter were more rapidly degraded during particle sinking [57].
This ratio was calculated also in the north Tyrrhenian Sea [65]. In the Central Tyrrhenian Sea higher LAP/GLU ratios in the bathypelagic layer than in the other ones were reported, suggesting high metabolic potentialities of prokaryotes in the degradation of proteinaceous material in the deep realm [63]. Also Monticelli et al. [58] observed high values of this ratio in a frontal system in the southern Adriatic Sea (Table 1).
In marine agammaegates different enzymatic ratios and an imbalance in bacterial enzymatic activity, with high values of protease activity and relatively low values of glucosidase activity, resulted in a progressive increase in the refractory component of the organic matter and a consequent increase in its C/N ratio [78]. The incomplete hydrolysis of organic matter related to low enzyme specificity lead to increasing recalcitrance and accumulation of long-lived organic matter, which could agammaegate into mucilage [70].
In the brackish waters of Ganzirri lake the decomposition of polysaccharides was enhanced during post-bloom and the LAP/GLU ratio increased from 1.18 in summer–autumn period to 7.17 during winter–spring [75].
The molar ratio between AP and LAP was proposed by Sala et al [48] to assess nutrient limitation in microbial population, being its values high when P and N are unbalanced.
AP/LAP ratios measured in the Ionian Sea during our previous studies showed different distribution patterns: a prevalence of AP over LAP and GLU was observed in the epipelagic layer and LAP/AP ratio was lower than 1, thus indicating that that microbial community was P-limited in surface layer (Table 1). In autumn 2000 similar low values were measured in all layers [46], suggesting that AP supplied the P necessary for protein synthesis, cell production and energy transport for both bacteria and phytoplankton [40],[71].
In the West and East Mediterranean waters the LAP/AP ratios were lower than 1 at surface during summer, indicating the quick remineralization of organic phosphates with prompt recycling of P in this layer [49]. In the surface layer of oligotrophic areas, a low ratio between LAP and AP, as proxy of P-limitation, suggested that bacteria are P limited, while the relative importance of AP decreased with increasing depth (Table 1). Also during spring and winter the needs of P for the primary production increased, so the LAP/AP ratio decreased [46],[58]. On the contrary in the Tyrrhenian waters the LAP/AP ratios showed values >1 both at surface and at high depths [63]. Deep waters were rarely P limited (Table 1), with some exceptions [55] where P limitation was found in both the deep and benthic boundary layers (LAP/AP = 0.4 and 0.6, respectively).
In the north Adriatic Sea, seasonal differences in the AP activity with high values in summer period were observed by Ivancic et al. [37] in both waters and agammaegates. In a seasonal study, AP prevailed over LAP during winter (LAP/AP = 0.09) suggesting that P was more efficiently recycled in this season than in summer; in winter, the high AP activity contributed to P needs supporting prokaryotic growth [40].
Also in the DCM a rapid recycling of P with respect to N and C was found in the Augusta bay [79].
The Mediterranean Sea offers then an attractive marine environment to study general ubiquitous processes across multiscale and multidirectional physical, biological and trophic gradients. Despite the numerous investigations of the last decades, the emerging picture of enzyme dynamics in this ecosystem is far from being satisfying, both at spatial and temporal scales. Some areas, especially in both the West and East Mediterranean basins, are still insufficiently known. More information is available in the Central Mediterranean Sea, where the relationship with temperature changes is evident [26]. At the epipelagic layers and in coastal zones an increase in the decomposition processes has been detected.
The temporal variability at seasonal and interannual scale also needs more intensive sampling and long-term records to investigate seasonal patterns and responses of the planktonic microbial metabolism to anthropogenic and climatic changes.
Although the relationships between climate and microbial processes need to be elucidated yet [27], in a warmer ocean microbial processes are expected to play an even more important role in the carbon cycle. In the context of climate changes, the fate of bacterial metabolism will depend on the rates at which labile organic carbon derived from primary producers will become available to incorporation into prokaryotic cells. However, the prediction of metabolic rates and the partitioning of their carbon uptake towards autotrophy or respiration by heterotrophs remain uncertain [18]. Long term studies on microbial activity and abundances with rising temperatures are needed to describe the evolutionary scenario and future biogeochemical implications of microbial processes in the Mediterranean Sea.
| [1] |
Jun SH, Hirata A, Kanai T, et al. (2014) The X-ray crystal structure of the euryarchaeal RNA polymerase in an open-clamp configuration. Nat Commun 5: 5132. doi: 10.1038/ncomms6132

|
| [2] |
Leśniewska E, Boguta M (2017) Novel layers of RNA polymerase III control affecting tRNA gene transcription in eukaryotes. Open Biol 7: 170001. doi: 10.1098/rsob.170001
|
| [3] | Torreira E, Louro JA, Pazos I, et al. (2017) The dynamic assembly of distinct RNA polymerase I complexes modulates rDNA transcription. Elife, 6. |
| [4] |
Werner F, Grohmann D (2011) Evolution of multisubunit RNA polymerases in the three domains of life. Nat Rev Microbiol 9: 85–98. doi: 10.1038/nrmicro2507
|
| [5] |
Werner M, Thuriaux P, Soutourina J (2009) Structure-function analysis of RNA polymerases I and III. Curr Opin Struct Biol 19: 740–745. doi: 10.1016/j.sbi.2009.10.005
|
| [6] |
Khatter H, Vorländer MK, Müller CW (2017) RNA polymerase I and III: Similar yet unique. Curr Opin Struct Biol 47: 88–94. doi: 10.1016/j.sbi.2017.05.008
|
| [7] |
Fong N, Saldi T, Sheridan RM, et al. (2017) RNA Pol II Dynamics Modulate Co-transcriptional Chromatin Modification, CTD Phosphorylation, and Transcriptional Direction. Mol Cell 66: 546. doi: 10.1016/j.molcel.2017.04.016
|
| [8] |
Haag JR, Pikaard CS (2011) Multisubunit RNA polymerases IV and V: Purveyors of non-coding RNA for plant gene silencing. Nat Rev Mol Cell Biol1 12: 483–492. doi: 10.1038/nrm3152
|
| [9] |
Böhmdorfer G, Rowley MJ, Kuciński J, et al. (2014) RNA‐directed DNA methylation requires stepwise binding of silencing factors to long non-coding RNA. Plant J Cell Mol Biol 79: 181–191. doi: 10.1111/tpj.12563
|
| [10] |
Haag JR, Brower-Toland B, Krieger EK, et al. (2014) Functional diversification of maize RNA polymerase IV and V subtypes via alternative catalytic subunits. Cell Rep 9: 378–390. doi: 10.1016/j.celrep.2014.08.067
|
| [11] |
Bernecky C, Herzog F, Baumeister W, et al. (2016) Structure of transcribing mammalian RNA polymerase II. Nature 529: 551–554. doi: 10.1038/nature16482
|
| [12] |
Kwapisz M, Beckouët F, Thuriaux P (2008) Early evolution of eukaryotic DNA-dependent RNA polymerases. Trends Genet 24: 211–215. doi: 10.1016/j.tig.2008.02.002
|
| [13] |
Zaros C, Briand JF, Boulard Y, et al. (2007) Functional organization of the Rpb5 subunit shared by the three yeast RNA polymerases. Nucleic Acids Res 35: 634–647. doi: 10.1093/nar/gkl686
|
| [14] |
Woychik NA, Liao SM, Kolodziej PA, et al. (1990) Subunits shared by eukaryotic nuclear RNA polymerases. Genes Dev 4: 313–323. doi: 10.1101/gad.4.3.313
|
| [15] |
Shpakovski GV, Acker J, Wintzerith M, et al. (1995) Four subunits that are shared by the three classes of RNA polymerase are functionally interchangeable between Homo sapiens and Saccharomyces cerevisiae. Mol Cell Biol 15: 4702–4710. doi: 10.1128/MCB.15.9.4702
|
| [16] | Kelly S, Wickstead B, Gull K (2005) An in silico analysis of trypanosomatid RNA polymerases: Insights into their unusual transcription. Biochem Soc Trans 33: 1435–1437. |
| [17] |
Armache KJ, Mitterweger S, Meinhart A, et al. (2005) Structures of complete RNA polymerase II and its subcomplex, Rpb4/7. J Biol Chem 280: 7131–7134. doi: 10.1074/jbc.M413038200
|
| [18] | Bushnell DA, Kornberg RD (2003) Complete, 12-subunit RNA polymerase II at 4.1-A resolution: Implications for the initiation of transcription. Proc Natl Acad Sci U S A 100: 6969–6973. |
| [19] | Cramer P, Bushnell DA, Kornberg RD (2001) Structural basis of transcription: RNA polymerase II at 2.8 Ångstrom resolution. Science 292: 1863–1876. |
| [20] |
Fernándeztornero C, Morenomorcillo M, Rashid UJ, et al. (2013) Crystal structure of the 14-subunit RNA polymerase I. Nature 502: 644–649. doi: 10.1038/nature12636
|
| [21] |
Hoffmann NA, Jakobi AJ, Maria MM, et al. (2015) Molecular structures of unbound and transcribing RNA polymerase III. Nature 528: 231–236. doi: 10.1038/nature16143
|
| [22] |
Nonet M, Scafe C, Sexton J, et al. (1987) Eucaryotic RNA polymerase conditional mutant that rapidly ceases mRNA synthesis. Mol Cell Biol 7: 1602–1611. doi: 10.1128/MCB.7.5.1602
|
| [23] |
Mitsuzawa H, Ishihama A (2004) RNA polymerase II transcription apparatus in Schizosaccharomyces pombe. Curr Genet 44: 287–294. doi: 10.1007/s00294-003-0446-8
|
| [24] |
Langer D, Hain J, Thuriaux P, et al. (1995) Transcription in archaea: Similarity to that in eucarya. Proc Natl Acad Sci U S A 92: 5768–5772. doi: 10.1073/pnas.92.13.5768
|
| [25] |
Korkhin Y, Unligil UM, Littlefield O, et al. (2009) Evolution of complex RNA polymerases: The complete archaeal RNA polymerase structure. PLoS Biol 7: e1000102. doi: 10.1371/journal.pbio.1000102
|
| [26] |
Sommer B, Waege I, Pöllmann D, et al. (2014) Activation of a chimeric Rpb5/RpoH subunit using library selection. PLoS One 9: e87485. doi: 10.1371/journal.pone.0087485
|
| [27] |
Iyer LM, Balaji S, Koonin EV, et al. (2006) Evolutionary genomics of nucleo-cytoplasmic large DNA viruses. Virus Res 117: 156–184. doi: 10.1016/j.virusres.2006.01.009
|
| [28] | Raoult D, Audic S, Robert C, et al. (2004) The 1.2-megabase genome sequence of Mimivirus. Science 306: 1344–1350. |
| [29] |
Cramer P, Bushnell DA, Fu J, et al. (2000) Architecture of RNA polymerase II and implications for the transcription mechanism. Science 288: 640–649. doi: 10.1126/science.288.5466.640
|
| [30] |
Todone F, Brick P, Onesti S (2000) Crystal structure of RPB5, a universal eukaryotic RNA polymerase subunit and transcription factor interaction target. Proc Natl Acad Sci U S A 97: 6306–6310. doi: 10.1073/pnas.97.12.6306
|
| [31] |
Navarro F, Thuriaux P (2000) In vivo misreading by tRNA overdose. RNA 6: 103–110. doi: 10.1017/S1355838200991714
|
| [32] |
Flores A, Briand JF, Gadal O, et al. (1999) A protein-protein interaction map of yeast RNA polymerase III. Proc Natl Acad Sci U S A 96: 7815–7820. doi: 10.1073/pnas.96.14.7815
|
| [33] |
Grünberg S, Reich C, Zeller ME, et al. (2010) Rearrangement of the RNA polymerase subunit H and the lower jaw in archaeal elongation complexes. Nucleic Acids Res 38: 1950–1963. doi: 10.1093/nar/gkp1190
|
| [34] |
Barnes CO, Calero M, Malik I, et al. (2015) Crystal structure of a transcribing RNA polymerase II complex reveals a complete transcription bubble. Mol Cell 59: 258–269. doi: 10.1016/j.molcel.2015.06.034
|
| [35] |
Kimura M, Ishiguro A, Ishihama A (1997) RNA polymerase II subunits 2, 3, and 11 form a core subassembly with DNA binding activity. J Biol Chem 272: 25851–25855. doi: 10.1074/jbc.272.41.25851
|
| [36] |
Bartholomew B, Durkovich D, Kassavetis G, et al. (1993) Orientation and topography of RNA polymerase III in transcription complexes. Mol Cell Biol 13: 942–952. doi: 10.1128/MCB.13.2.942
|
| [37] |
He Y, Fang J, Taatjes DJ, et al. (2013) Structural visualization of key steps in human transcription initiation. Nature 495: 481. doi: 10.1038/nature11991
|
| [38] |
Fishburn J, Tomko E, Galburt E, et al. (2015) Double-stranded DNA translocase activity of transcription factor TFIIH and the mechanism of RNA polymerase II open complex formation. Proc Natl Acad Sci U S A 112: 3961–3966. doi: 10.1073/pnas.1417709112
|
| [39] |
Soutourina J, Bordas-Le FV, Gendrel G, et al. (2006) Rsc4 connects the chromatin remodeler RSC to RNA polymerases. Mol Cell Biol 26: 4920–4933. doi: 10.1128/MCB.00415-06
|
| [40] |
Peeters E, Driessen RP, Werner F, et al. (2015) The interplay between nucleoid organization and transcription in archaeal genomes. Nat Rev Microbiol 13: 333. doi: 10.1038/nrmicro3467
|
| [41] |
Miyao T, Woychik NA (1998) RNA polymerase subunit RPB5 plays a role in transcriptional activation. Proc Natl Acad Sci U S A 95: 15281–15286. doi: 10.1073/pnas.95.26.15281
|
| [42] | Cheong JH, Yi M, Lin Y, et al. (1995) Human RPB5, a subunit shared by eukaryotic nuclear RNA polymerases, binds human hepatitis B virus X protein and may play a role in X transactivation. EMBO J 14: 143–150. |
| [43] |
Haviv I, Shamay MG, Shaul Y (1998) Hepatitis B virus pX targets TFIIB in transcription coactivation. Mol Cell Biol 18: 1562–1569. doi: 10.1128/MCB.18.3.1562
|
| [44] |
Dorjsuren D, Lin Y, Wei W, et al. (1998) RMP, a novel RNA polymerase II subunit 5-interacting protein, counteracts transactivation by hepatitis B virus X protein. Mol Cell Biol 18: 7546–7555. doi: 10.1128/MCB.18.12.7546
|
| [45] |
Le TT, Zhang S, Hayashi N, et al. (2005) Mutational analysis of human RNA polymerase II subunit 5 (RPB5): The residues critical for interactions with TFIIF subunit RAP30 and hepatitis B virus X protein. J Biochem 138: 215–224. doi: 10.1093/jb/mvi119
|
| [46] |
Wei W, Dorjsuren D, Lin Y, et al. (2001) Direct interaction between the subunit RAP30 of transcription factor IIF (TFIIF) and RNA polymerase subunit 5, which contributes to the association between TFIIF and RNA polymerase II. J Biol Chem 276: 12266–12273. doi: 10.1074/jbc.M009634200
|
| [47] |
Wei W, Gu JX, Zhu CQ, et al. (2003) Interaction with general transcription factor IIF (TFIIF) is required for the suppression of activated transcription by RPB5-mediating protein (RMP). Cell Res 13: 111. doi: 10.1038/sj.cr.7290155
|
| [48] |
Morohoshi F, Arai K, Takahashi EI, et al. (1996) Cloning and Mapping of a Human RBP56 Gene Encoding a Putative RNA Binding Protein Similar to FUS/TLS and EWS Proteins. Genomics 38: 51–57. doi: 10.1006/geno.1996.0591
|
| [49] | Bertolotti A, Lutz Y, Heard DJ, et al. (1996) HTAF(II)68, a novel RNA/ssDNA-binding protein with homology to the pro-oncoproteins TLS/FUS and EWS is associated with both TFIID and RNA polymerase II. EMBO J 15: 5022. |
| [50] |
Bertolotti A, Melot T, Acker J, et al. (1998) EWS, but not EWS-FLI-1, is associated with both TFIID and RNA polymerase II: Interactions between two members of the TET family, EWS and hTAFII68, and subunits of TFIID and RNA polymerase II complexes. Mol Cell Biol 18: 1489–1497. doi: 10.1128/MCB.18.3.1489
|
| [51] |
Makino Y, Yogosawa S, Kayukawa K, et al. (1999) TATA-binding protein-interacting protein 120, TIP120, stimulates three classes of eukaryotic transcription via a unique mechanism. Mol Cell Biol 19: 7951–7960. doi: 10.1128/MCB.19.12.7951
|
| [52] |
Grünberg S, Hahn S (2013) Structural insights into transcription initiation by RNA polymerase II. Trends Biochem Sci 38: 603–611. doi: 10.1016/j.tibs.2013.09.002
|
| [53] |
Martínezfernández V, Garridogodino AI, Miróngarcía MC, et al. (2017) Rpb5 modulates the RNA polymerase II transition from initiation to elongation by influencing Spt5 association and backtracking. Biochim Biophys Acta 1861: 1–13. doi: 10.1016/j.bbagen.2016.11.023
|
| [54] | Martínezfernández V, Garridogodino AI, Cuevas-Bermudez A, et al. (2015) Cytoplasmic and Nuclear Functions for the Prefoldin-like URI/Bud27. Nova Science Publishers. |
| [55] |
Miróngarcía MC, Garridogodino AI, Garcíamolinero V, et al. (2013) The prefoldin Bud27 mediates the assembly of the eukaryotic RNA polymerases in an Rpb5-dependent manner. PLoS Genet 9: e1003297. doi: 10.1371/journal.pgen.1003297
|
| [56] |
Gstaiger M, Luke B, Hess D, et al. (2003) Control of nutrient-sensitive transcription programs by the unconventional prefoldin URI. Science 302: 1208–1212. doi: 10.1126/science.1088401
|
| [57] |
Riveracalzada A, Pal M, Muñozhernández H, et al. (2017) The structure of the R2TP complex defines a platform for recruiting diverse client proteins to the HSP90 molecular chaperone system. Structure 25: 1145–1152. doi: 10.1016/j.str.2017.05.016
|
| [58] |
Boulon S, Bertrand E, Pradet-Balade B (2012) HSP90 and the R2TP co-chaperone complex: Building multi-protein machineries essential for cell growth and gene expression. RNA Biolo 9: 148–154. doi: 10.4161/rna.18494
|
| [59] |
Boulon S, Pradet-Balade B, Verheggen C, et al. (2010) HSP90 and its R2TP/Prefoldin-like cochaperone are involved in the cytoplasmic assembly of RNA polymerase II. Mol Cell 39: 912–924. doi: 10.1016/j.molcel.2010.08.023
|
| [60] |
Mita P, Savas JN, Ha S, et al. (2013) Analysis of URI nuclear interaction with RPB5 and components of the R2TP/Prefoldin-like complex. PLoS One 8: e63879. doi: 10.1371/journal.pone.0063879
|
| [61] |
Cloutier P, Poitras C, Durand M, et al. (2017) R2TP/Prefoldin-like component RUVBL1/RUVBL2 directly interacts with ZNHIT2 to regulate assembly of U5 small nuclear ribonucleoprotein. Nat Commun 8: 15615. doi: 10.1038/ncomms15615
|
| [62] |
Yart A, Gstaiger M, Wirbelauer C, et al. (2005) The HRPT2 tumor suppressor gene product parafibromin associates with human PAF1 and RNA polymerase II. Mol Cell Biol 25: 5052–5060. doi: 10.1128/MCB.25.12.5052-5060.2005
|
| [63] | Parusel CT, Kritikou EA, Hengartner MO, et al. (2006) URI-1 is required for DNA stability in C. elegans. Development 133: 621–629. |
| [64] |
Kirchner J, Vissi E, Gross S, et al. (2008) Drosophila Uri, a PP1alpha binding protein, is essential for viability, maintenance of DNA integrity and normal transcriptional activity. BMC Mol Biol 9: 1–17. doi: 10.1186/1471-2199-9-1
|
| [65] |
Tummala KS, Gomes AL, Yilmaz M, et al. (2014) Inhibition of De Novo NAD Synthesis by Oncogenic URI Causes Liver Tumorigenesis through DNA Damage. Cancer Cell 26: 826–839. doi: 10.1016/j.ccell.2014.10.002
|
| [66] |
Burén S, Gomes AL, Teijeiro A, et al. (2016) Regulation of OGT by URI in Response to Glucose Confers c-MYC-Dependent Survival Mechanisms. Cancer Cell 30: 290–307. doi: 10.1016/j.ccell.2016.06.023
|
| [67] |
Deplazes A, Mockli N, Luke B, et al. (2009) Yeast Uri1p promotes translation initiation and may provide a link to cotranslational quality control. EMBO J 28: 1429–1441. doi: 10.1038/emboj.2009.98
|
| [68] |
Lin Y, Nomura T, Cheong J, et al. (1997) Hepatitis B virus X protein is a transcriptional modulator that communicates with transcription factor IIB and the RNA polymerase II subunit 5. J Biol Chem 272: 7132–7139. doi: 10.1074/jbc.272.11.7132
|
| [69] |
Mita P, Savas JN, Djouder N, et al. (2011) Regulation of androgen receptor-mediated transcription by RPB5 binding protein URI/RMP. Mol Cell Biol 31: 3639–3652. doi: 10.1128/MCB.05429-11
|
| [70] |
Vernekar DV, Bhargava P (2015) Yeast Bud27 modulates the biogenesis of Rpc128 and Rpc160 subunits and the assembly of RNA polymerase III. Biochim Biophys Acta 1849: 1340–1353. doi: 10.1016/j.bbagrm.2015.09.010
|
| [71] |
Ciesla M, Makala E, Plonka M, et al. (2015) Rbs1, a new protein implicated in RNA polymerase III biogenesis in yeast Saccharomyces cerevisiae. Mol Cell Biol 35: 1169–1181. doi: 10.1128/MCB.01230-14
|
| [72] |
Miróngarcía MC, Garridogodino AI, Martínezfernández V, et al. (2014) The yeast prefoldin-like URI-orthologue Bud27 associates with the RSC nucleosome remodeler and modulates transcription. Nucleic Acids Res 42: 9666–9676. doi: 10.1093/nar/gku685
|
| 1. | Alice Madonia, Gabriella Caruso, Daniele Piazzolla, Simone Bonamano, Viviana Piermattei, Giuseppe Zappalà, Marco Marcelli, Chromophoric Dissolved Organic Matter as a Tracer of Fecal Contamination for Bathing Water Quality Monitoring in the Northern Tyrrhenian Sea (Latium, Italy), 2020, 8, 2077-1312, 430, 10.3390/jmse8060430 | |
| 2. | Renata Zaccone, Cristina Misic, Filippo Azzaro, Maurizio Azzaro, Giovanna Maimone, Olga Mangoni, Giannetta Fusco, Alessandro Rappazzo, Rosabruna La Ferla, Regulation of Microbial Activity Rates by Organic Matter in the Ross Sea during the Austral Summer 2017, 2020, 8, 2076-2607, 1273, 10.3390/microorganisms8091273 | |
| 3. | Vincenzo Manna, Cinzia De Vittor, Michele Giani, Paola Del Negro, Mauro Celussi, Long-term patterns and drivers of microbial organic matter utilization in the northernmost basin of the Mediterranean Sea, 2021, 164, 01411136, 105245, 10.1016/j.marenvres.2020.105245 | |
| 4. | Marta Zdanowicz, Piotr Perliński, Zbigniew J. Mudryk, Extracellular hydrolytic enzyme activity in the water from the ponds of a carp farm, 2021, 76, 0006-3088, 1063, 10.2478/s11756-020-00659-9 | |
| 5. | France Van Wambeke, Elvira Pulido, Philippe Catala, Julie Dinasquet, Kahina Djaoudi, Anja Engel, Marc Garel, Sophie Guasco, Barbara Marie, Sandra Nunige, Vincent Taillandier, Birthe Zäncker, Christian Tamburini, Spatial patterns of ectoenzymatic kinetics in relation to biogeochemical properties in the Mediterranean Sea and the concentration of the fluorogenic substrate used, 2021, 18, 1726-4189, 2301, 10.5194/bg-18-2301-2021 | |
| 6. | Franco Decembrini, Carmela Caroppo, Gabriella Caruso, Alessandro Bergamasco, Linking Microbial Functioning and Trophic Pathways to Ecological Status in a Coastal Mediterranean Ecosystem, 2021, 13, 2073-4441, 1325, 10.3390/w13091325 | |
| 7. | Judit Boadella, Andrea Butturini, Jordi Compte, Giulia Gionchetta, Núria Perujo, Xavier D. Quintana, Anna M. Romaní, Different microbial functioning in natural versus man-made Mediterranean coastal lagoons in relation to season, 2021, 259, 02727714, 107434, 10.1016/j.ecss.2021.107434 | |
| 8. | Maria Papale, Gabriella Caruso, Giovanna Maimone, Rosabruna La Ferla, Angelina Lo Giudice, Alessandro Ciro Rappazzo, Alessandro Cosenza, Filippo Azzaro, Roberta Ferretti, Rodolfo Paranhos, Anderson Souza Cabral, Massimo Caccia, Angelo Odetti, Giuseppe Zappalà, Gabriele Bruzzone, Maurizio Azzaro, Microbial Community Abundance and Metabolism Close to the Ice-Water Interface of the Blomstrandbreen Glacier (Kongsfjorden, Svalbard): A Sampling Survey Using an Unmanned Autonomous Vehicle, 2023, 15, 2073-4441, 556, 10.3390/w15030556 | |
| 9. | Ingrid Ivančić, Romina Kraus, Mirjana Najdek, Stefano Cozzi, Ecological Importance of Alkaline Phosphatase Activity in Changing Marine Environmental Conditions, 2021, 13, 2073-4441, 2750, 10.3390/w13192750 | |
| 10. | Loredana Stabili, Lucia Rizzo, Rosa Caprioli, Giorgio Alabiso, Stefano Piraino, How Much Recurrent Outbreaks of the Moon Jellyfish May Impact the Dynamics of Bacterial Assemblages in Coastal Lagoons?, 2022, 14, 2073-4441, 3908, 10.3390/w14233908 | |
| 11. | Marta Małecka-Adamowicz, Łukasz Kubera, Patterns of Structural and Functional Bacterioplankton Metacommunity along a River under Anthropogenic Pressure, 2021, 13, 2071-1050, 11518, 10.3390/su132011518 | |
| 12. | Carmela Caroppo, Filippo Azzaro, Alessandro Bergamasco, Gabriella Caruso, Franco Decembrini, Phytoplankton and Bacterial Communities’ Patterns in a Highly Dynamic Ecosystem (Central Mediterranean Sea), 2022, 14, 2073-4441, 2057, 10.3390/w14132057 | |
| 13. | Gabriella Caruso, Maria Grazia Giacobbe, Filippo Azzaro, Franco Decembrini, Marcella Leonardi, Stefano Miserocchi, Xiuyun Cao, Chunlei Song, Yiyong Zhou, All-In-One: Microbial Response to Natural and Anthropogenic Forcings in a Coastal Mediterranean Ecosystem, the Syracuse Bay (Ionian Sea, Italy), 2021, 10, 2077-1312, 19, 10.3390/jmse10010019 | |
| 14. | Zahra Akbari Ariadji, Gede Suantika, Pingkan Aditiawati, Trophic state assessment of marine ecosystems used for floating net cage aquaculture: a case study in Pegametan Bay, North Bali, Indonesia, 2024, 32, 0967-6120, 6611, 10.1007/s10499-024-01481-9 | |
| 15. | Ke-Xuan Huang, Yu-Xuan Jiang, Yan-Ru Dang, Qi-Long Qin, Release of intracellular enzymes increases the total extracellular activities of carbohydrate-processing enzymes in marine environment, 2024, 371, 1574-6968, 10.1093/femsle/fnae077 | |
| 16. | Zhenbo Chen, Zhihong Liu, Changchun Song, Agricultural fertilization near marshes impacts the potential for greenhouse gas emissions from wetland ecosystems by modifying microbial communities, 2024, 930, 00489697, 172400, 10.1016/j.scitotenv.2024.172400 | |
| 17. | Simone Cappello, Francesco Smedile, Gabriella Caruso, Sabrina Patania, Alessia Lunetta, Maurizio Azzaro, A Snapshot of the Taxonomic Composition and Metabolic Activity of the Microbial Community in an Arctic Harbour (Ny-Ålesund, Kongsfjorden, Svalbard), 2023, 11, 2077-1312, 1018, 10.3390/jmse11051018 | |
| 18. | Yulia Polyak, Yulia Gubelit, Lyudmila Bakina, Tatyana Shigaeva, Valentina Kudryavtseva, Impact of macroalgal blooms on biogeochemical processes in estuarine systems: a case study in the eastern Gulf of Finland, Baltic Sea, 2024, 24, 1439-0108, 1854, 10.1007/s11368-023-03630-8 | |
| 19. | Xiyin Huang, Qiong Tang, Siqin Liu, Chen Li, Yaoguang Li, Yunjun Sun, Xuezhi Ding, Liqiu Xia, Shengbiao Hu, Discovery of an antitumor compound from xenorhabdus stockiae HN_xs01, 2024, 40, 0959-3993, 10.1007/s11274-024-03915-1 | |
| 20. | Carmen Rizzo, Erika Arcadi, Rosario Calogero, Alessandro Ciro Rappazzo, Gabriella Caruso, Giovanna Maimone, Angelina Lo Giudice, Franco Andaloro, Teresa Romeo, 2024, Microcosm Culture of a Shallow Hydrothermal Water Reveals a Shift of Microbial Communities, 979-8-3503-7900-6, 1, 10.1109/MetroSea62823.2024.10765722 | |
| 21. | Lai Wei, Ya Yang, Hongwen Gao, Rui Wang, Feng Cao, Qinghui Huang, Biogeochemical impact on the distribution variations of organophosphorus flame retardants in estuarine area and insight into climate change, 2025, 491, 03043894, 137993, 10.1016/j.jhazmat.2025.137993 |
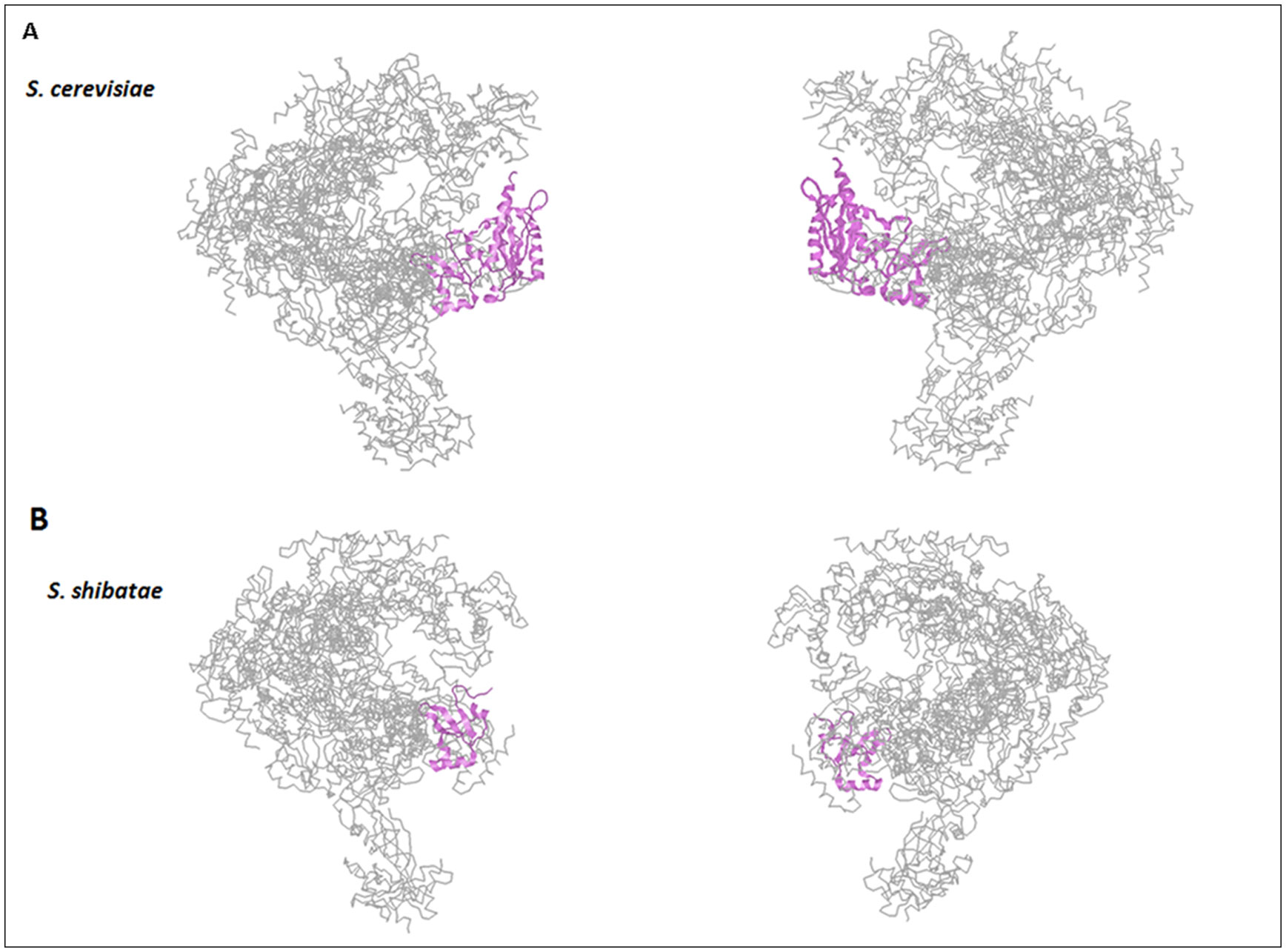
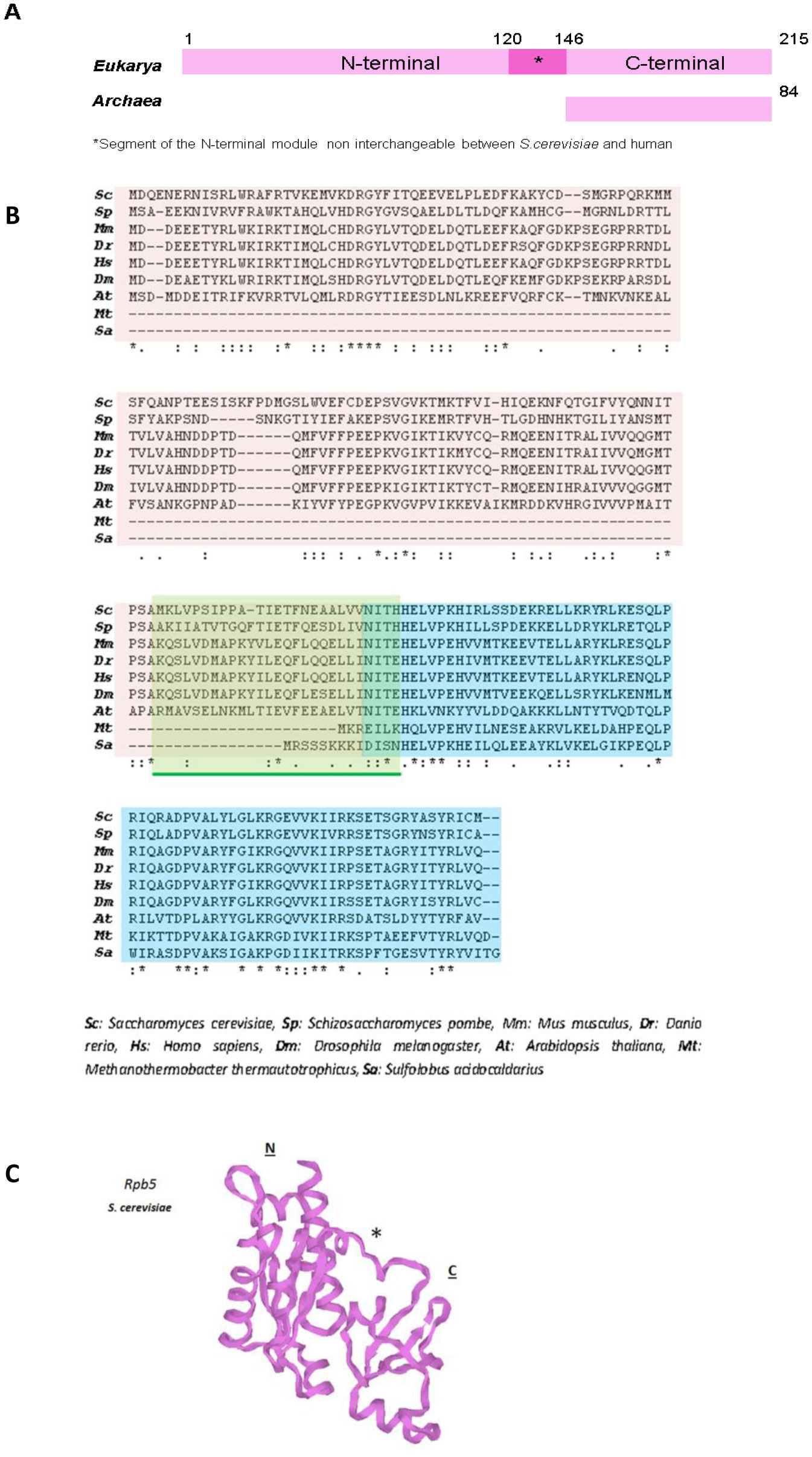
Figures(2) / Tables(2)

Verónica Martínez-Fernández, Francisco Navarro. Rpb5, a subunit shared by eukaryotic RNA polymerases, cooperates with prefoldin-like Bud27/URI[J]. AIMS Genetics, 2018, 5(1): 63-74. doi: 10.3934/genet.2018.1.63

| Area | Depth (m) | Sites | season | T °C | S | LAP(nM h−1)* | GLU(nM h−1)* | AP(nM h−1)* | LAP/GLU molar ratio | LAP/AP molar ratio | References |
| NW Mediterranean | 10–2000 | DYFAMED area | spring | >12.70 | 38.40 | 1.78–0.23 | 1.12–0.22 | 1.6 | [61] | ||
| 10–2000 | DYFAMED area | fall | 1.44–0.45 | 1.5–1.2 | 0.37 | " | |||||
| NW Mediterranean | DYFAMED area | autumn 04 | 3.7–4.6 | 2.4–3.2 | [53] | ||||||
| Mediterranean | 5–200 | W-E transect | summer 99 | 22.7–24.9 | 37.28–39.01 | 1.1–9.3 | 0.62–1.65 | [51] | |||
| NW Mediterranean | 5 | Marseille Bay | annual 12-13 | 12.8–25.8 | 38.15 | 5.1–85.1 | 0.1–115.7 | 5.42 | [71] | ||
| Ligurian Sea | 5–200 | gulf of Genoa | summer 02 | 17–24 | >36.7 | 1.78–4.04 | 0.04–0.19 | 1.52 | 25-68 | [68] | |
| Ligurian Sea | 0.5 | Portofino | spring -summer 07 | 19.4 | 110.1–165.6 | 13.9–29.4 | [66] | ||||
| South Tyrrhenian Sea | 1–100 | Eolian Island | spring 04 | 1.9–2.5 | 0.04–0.08 | [65] | |||||
| Tyrrhenian sea | 25–3000 | winter 05 | 13–16 | 37.8–38.25 | 0.3–21.7 | 0.5–9.9 | [62] | ||||
| South Tyrrhenian Sea | 5–100 | summer 05 | 17.55 | 0.59 | 3.60 | 29.75 | 4.88 | [63] | |||
| " | 200–1000 | " | 23.71 | 0.80 | 0.78 | 29.64 | 30.40 | " | |||
| " | >1000 | " | 37.72 | 0.40 | 0.93 | 94.3 | 40.56 | " | |||
| Tyrrhenian sea | 10–200 | st. VECTOR | summer 07 | 16.79 | 38.08 | 0.31 | 0.31 | 1.16 | 1 | 0.27 | [49] |
| " | 500–1000 | " | " | 13.81 | 38.69 | 0.37 | 0.37 | 0.19 | 1 | 1.95 | " |
| " | 1500–3500 | " | " | 13.49 | 38.53 | 0.61 | 0.47 | 0.20 | 1.29 | 3.05 | " |
| Ionian Sea | 5–200 | pelagic | winter 98 | 15.17 | 38.59 | 1.92 | 0.19 | 1.30 | 9.91 | 0.67 | [55] |
| 400–1000 | " | 13.98 | 38.80 | 1.59 | 0.16 | 0.16 | 9.94 | 0.10 | " | ||
| 1100-4000 | pelagic | " | 13.92 | 38.76 | 0.51 | 0.06 | 0.92 | 8.81 | 1.79 | " | |
| Ionian Sea | 5–200 | pelagic | winter 99 | 15.79 | 38.65 | 5.40 | 4.85 | 9.97 | 1.12 | 0.54 | [26], [46] |
| 400–1000 | " | 13.60 | 38.73 | 2.83 | 0,70 | 1.90 | 4.05 | 1.49 | " | ||
| 2000–3300 | " | 13.39 | 38.71 | 5.97 | 0.49 | 11.97 | 12.17 | 0.50 | " | ||
| Ionian Sea | 5–200 | pelagic | autumn 00 | 17.47 | 38.68 | 1,48 | 1,79 | 99.86 | 0.83 | 0.02 | [26],[46] |
| 400–1000 | " | 13.77 | 38,76 | 1,58 | 0,51 | 52.27 | 3.11 | 0.30 | " | ||
| 2000–3300 | " | 13.79 | 38.73 | 1,53 | 2,86 | 35.41 | 0.53 | 0.04 | " | ||
| Ionian Sea | 5–200 | pelagic | summer 01 | 17.38 | 38.62 | 2,02 | 0,20 | 5,08 | 10.12 | 0.40 | [26],[46] |
| 400–1000 | " | 13.81 | 38.76 | 10,64 | 0,35 | 4,82 | 29.97 | 2.21 | " | ||
| 2000–3300 | " | 13.79 | 38.73 | 1,08 | 0,34 | 3,12 | 3.17 | 0.35 | " | ||
| Ionian Sea | 5–200 | pelagic area | autumn 04 | 17.14 | 38.44 | 4,95 | 2,04 | 4,06 | 2,43 | 1,22 | [26],[46] |
| 500–1000 | " | 13.96 | 38.80 | 3,60 | 0,34 | 1,63 | 10.46 | 2,21 | " | ||
| 1100– 4000 | " | 13.85 | 38.80 | 4,53 | 0,80 | 2.01 | 5.67 | 2,25 | " | ||
| Ionian Sea | 5–200 | pelagic area | spring 02 | 15.49 | 38.58 | 2,96 | 1,44 | 5.29 | 2.05 | 0.56 | [26],[46] |
| 500–1000 | " | 14.02 | 38.79 | 0,50 | 0,10 | 4.87 | 0,97 | " | |||
| 1100– 4000 | " | 13.84 | 38.74 | 0,33 | 0,09 | 0,16 | 3.70 | 2,03 | " | ||
| Ionian Sea | 0–150 | pelagic area | autumn 11 | >15.50 | <38.80 | 21.10 | 18.91 | 144.99 | 1.12 | 0.15 | [64] |
| 200-600 | " | " | < 15.00 | >39.00 | 20.84 | 15.03 | 14.26 | 1.39 | 1.46 | " | |
| >3000 | " | " | 13.42 | >38.72 | 20.39 | 12.94 | 12.21 | 1.58 | 1.67 | " | |
| Ionian Sea | 5–100 | pelagic area | summer 12 | 18.16 | 38.29 | 4.91 | 1.37 | 2.04 | 3,59 | 2,40 | [26],[40] |
| Ionian Sea | 5–100 | " | winter 13 | 14.97 | 38.45 | 1.49 | 0.27 | 16.74 | 5.53 | 0.09 | " |
| Ionian Sea | 1–60 | coastal station | autumn 01 | 19.26 | 38.53 | 207.29 | 11.28 | 717.8 | 18.37 | 0,29 | [79] |
| Atlantic ocean | 5–100 | Atlantic station | summer 07 | 0.38 | 0.27 | 0.22 | 1.41 | 1.73 | [49] | ||
| 200–1000 | " | 0.74 | 0.13 | 0.05 | 5.69 | 14.8 | " | ||||
| 1500–3000 | " | 0.57 | 0.25 | 0.01 | 2.28 | 57 | " | ||||
| West Mediterranean | 5–200 | pelagic area | summer 07 | 16.21 | 37.8 | 0.35 | 0.20 | 1.21 | 1.75 | 0.29 | [49] |
| 500–1000 | " | 13.36 | 38.57 | 0.21 | 0.32 | 0.19 | 0.66 | 1,11 | " | ||
| West Mediterranean | 1500–3500 | " | 13.29 | 38.50 | 0.36 | 0.25 | 0.20 | 1.44 | 1.8 | " | |
| East Mediterranean | 5–200 | summer 07 | 18.41 | 38.79 | 1.53 | 0.50 | 4.56 | 3.06 | 0.33 | [49] | |
| 500–1000 | " | 14.01 | 38.81 | 0.66 | 0.52 | 0.33 | 1.27 | 2.01 | " | ||
| 1500– 2900 | " | 13.95 | 38.75 | 1.22 | 0.58 | 0.78 | 2.10 | 1.56 | " | ||
| West Mediterranean | 5–200 | 2.18 | 0.88 | 0.51 | [59] | ||||||
| 500–1000 | 1.04 | 0.51 | 0.35 | " | |||||||
| West Mediterranean | 1500–3500 | 0.66 | 0.41 | 0.31 | " | ||||||
| East Mediterranean | 5–200 | 0.08 | 0.31 | 0.20 | [59] | ||||||
| 500–1000 | 0.09 | 0.10 | 0.13 | " | |||||||
| 1500– 2900 | 0.05 | 0.13 | 0.03 | " | |||||||
| North Adriatic Sea | gen-60 | coastal station | seasonal study | 2.2–1701 | 0.3–114 | 0.8 -1770 | [70] | ||||
| North Adriatic Sea | 1–60 | coastal station | spring-summer | 115– 67.5 | 2.1–5.2 | 13.6 | 22.14 | 4.96 | [73] | ||
| North Adriatic Sea | autumn | 2.04–4.64 | [38] | ||||||||
| North Adriatic Sea | 1–10 | coastal station | spring-autumn | 70–4640 | [37] | ||||||
| South Adriatic Sea | 5–100 | convection site | seasonal study | 13.8–15.6 | 38.71–38.74 | 0.01–7.86 | 0.003–3.5 | 0.05–31.96 | [57] | ||
| 100–800 | " | 13.6–13.6 | 38.74–38.77 | 0.01–2.66 | 0.003–0.98 | 0.003–6.47 | " | ||||
| " | 800–1200 | " | 12.9–13.1 | 38.72–38.73 | 0.01–0.15 | 0.003–0.95 | 0.08–17.96 | " | |||
| South Adriatic Sea | 0–30 | Gulf of Manfredonia | spring 03 | 18.44 | 37.92 | 13.99 | 1.13 | 43.32 | 12.4 | 0.32 | [58] |
| 0–30 | offshore waters | " | 17.25 | 38.73 | 31.18 | 0.48 | 43,28 | 72.51 | 0.72 | " | |
| >30 | " | " | 14.11 | 38.70 | 19.45 | 0.26 | 23.49 | 75.97 | 0.83 | " | |
| North Atlantic | surface | 4.2–24.9 | [27] | ||||||||
| South Atlantic | surface | 11.5–151.4 | " | ||||||||
| Atlantic ocean | 100–4500 | 0.2–13.8 | 0.05–0.25 | 0.14–1.3 | [34] | ||||||
| subtropical Atlantic | 100–4500 | 0.6–9.2 | 0.02–0.07 | 0.04–3.7 | [67] |
DownLoad:
CSV
| Area | Depth (m) | Sites | season | T °C | S | LAP(nM h−1)* | GLU(nM h−1)* | AP(nM h−1)* | LAP/GLU molar ratio | LAP/AP molar ratio | References |
| NW Mediterranean | 10–2000 | DYFAMED area | spring | >12.70 | 38.40 | 1.78–0.23 | 1.12–0.22 | 1.6 | [61] | ||
| 10–2000 | DYFAMED area | fall | 1.44–0.45 | 1.5–1.2 | 0.37 | " | |||||
| NW Mediterranean | DYFAMED area | autumn 04 | 3.7–4.6 | 2.4–3.2 | [53] | ||||||
| Mediterranean | 5–200 | W-E transect | summer 99 | 22.7–24.9 | 37.28–39.01 | 1.1–9.3 | 0.62–1.65 | [51] | |||
| NW Mediterranean | 5 | Marseille Bay | annual 12-13 | 12.8–25.8 | 38.15 | 5.1–85.1 | 0.1–115.7 | 5.42 | [71] | ||
| Ligurian Sea | 5–200 | gulf of Genoa | summer 02 | 17–24 | >36.7 | 1.78–4.04 | 0.04–0.19 | 1.52 | 25-68 | [68] | |
| Ligurian Sea | 0.5 | Portofino | spring -summer 07 | 19.4 | 110.1–165.6 | 13.9–29.4 | [66] | ||||
| South Tyrrhenian Sea | 1–100 | Eolian Island | spring 04 | 1.9–2.5 | 0.04–0.08 | [65] | |||||
| Tyrrhenian sea | 25–3000 | winter 05 | 13–16 | 37.8–38.25 | 0.3–21.7 | 0.5–9.9 | [62] | ||||
| South Tyrrhenian Sea | 5–100 | summer 05 | 17.55 | 0.59 | 3.60 | 29.75 | 4.88 | [63] | |||
| " | 200–1000 | " | 23.71 | 0.80 | 0.78 | 29.64 | 30.40 | " | |||
| " | >1000 | " | 37.72 | 0.40 | 0.93 | 94.3 | 40.56 | " | |||
| Tyrrhenian sea | 10–200 | st. VECTOR | summer 07 | 16.79 | 38.08 | 0.31 | 0.31 | 1.16 | 1 | 0.27 | [49] |
| " | 500–1000 | " | " | 13.81 | 38.69 | 0.37 | 0.37 | 0.19 | 1 | 1.95 | " |
| " | 1500–3500 | " | " | 13.49 | 38.53 | 0.61 | 0.47 | 0.20 | 1.29 | 3.05 | " |
| Ionian Sea | 5–200 | pelagic | winter 98 | 15.17 | 38.59 | 1.92 | 0.19 | 1.30 | 9.91 | 0.67 | [55] |
| 400–1000 | " | 13.98 | 38.80 | 1.59 | 0.16 | 0.16 | 9.94 | 0.10 | " | ||
| 1100-4000 | pelagic | " | 13.92 | 38.76 | 0.51 | 0.06 | 0.92 | 8.81 | 1.79 | " | |
| Ionian Sea | 5–200 | pelagic | winter 99 | 15.79 | 38.65 | 5.40 | 4.85 | 9.97 | 1.12 | 0.54 | [26], [46] |
| 400–1000 | " | 13.60 | 38.73 | 2.83 | 0,70 | 1.90 | 4.05 | 1.49 | " | ||
| 2000–3300 | " | 13.39 | 38.71 | 5.97 | 0.49 | 11.97 | 12.17 | 0.50 | " | ||
| Ionian Sea | 5–200 | pelagic | autumn 00 | 17.47 | 38.68 | 1,48 | 1,79 | 99.86 | 0.83 | 0.02 | [26],[46] |
| 400–1000 | " | 13.77 | 38,76 | 1,58 | 0,51 | 52.27 | 3.11 | 0.30 | " | ||
| 2000–3300 | " | 13.79 | 38.73 | 1,53 | 2,86 | 35.41 | 0.53 | 0.04 | " | ||
| Ionian Sea | 5–200 | pelagic | summer 01 | 17.38 | 38.62 | 2,02 | 0,20 | 5,08 | 10.12 | 0.40 | [26],[46] |
| 400–1000 | " | 13.81 | 38.76 | 10,64 | 0,35 | 4,82 | 29.97 | 2.21 | " | ||
| 2000–3300 | " | 13.79 | 38.73 | 1,08 | 0,34 | 3,12 | 3.17 | 0.35 | " | ||
| Ionian Sea | 5–200 | pelagic area | autumn 04 | 17.14 | 38.44 | 4,95 | 2,04 | 4,06 | 2,43 | 1,22 | [26],[46] |
| 500–1000 | " | 13.96 | 38.80 | 3,60 | 0,34 | 1,63 | 10.46 | 2,21 | " | ||
| 1100– 4000 | " | 13.85 | 38.80 | 4,53 | 0,80 | 2.01 | 5.67 | 2,25 | " | ||
| Ionian Sea | 5–200 | pelagic area | spring 02 | 15.49 | 38.58 | 2,96 | 1,44 | 5.29 | 2.05 | 0.56 | [26],[46] |
| 500–1000 | " | 14.02 | 38.79 | 0,50 | 0,10 | 4.87 | 0,97 | " | |||
| 1100– 4000 | " | 13.84 | 38.74 | 0,33 | 0,09 | 0,16 | 3.70 | 2,03 | " | ||
| Ionian Sea | 0–150 | pelagic area | autumn 11 | >15.50 | <38.80 | 21.10 | 18.91 | 144.99 | 1.12 | 0.15 | [64] |
| 200-600 | " | " | < 15.00 | >39.00 | 20.84 | 15.03 | 14.26 | 1.39 | 1.46 | " | |
| >3000 | " | " | 13.42 | >38.72 | 20.39 | 12.94 | 12.21 | 1.58 | 1.67 | " | |
| Ionian Sea | 5–100 | pelagic area | summer 12 | 18.16 | 38.29 | 4.91 | 1.37 | 2.04 | 3,59 | 2,40 | [26],[40] |
| Ionian Sea | 5–100 | " | winter 13 | 14.97 | 38.45 | 1.49 | 0.27 | 16.74 | 5.53 | 0.09 | " |
| Ionian Sea | 1–60 | coastal station | autumn 01 | 19.26 | 38.53 | 207.29 | 11.28 | 717.8 | 18.37 | 0,29 | [79] |
| Atlantic ocean | 5–100 | Atlantic station | summer 07 | 0.38 | 0.27 | 0.22 | 1.41 | 1.73 | [49] | ||
| 200–1000 | " | 0.74 | 0.13 | 0.05 | 5.69 | 14.8 | " | ||||
| 1500–3000 | " | 0.57 | 0.25 | 0.01 | 2.28 | 57 | " | ||||
| West Mediterranean | 5–200 | pelagic area | summer 07 | 16.21 | 37.8 | 0.35 | 0.20 | 1.21 | 1.75 | 0.29 | [49] |
| 500–1000 | " | 13.36 | 38.57 | 0.21 | 0.32 | 0.19 | 0.66 | 1,11 | " | ||
| West Mediterranean | 1500–3500 | " | 13.29 | 38.50 | 0.36 | 0.25 | 0.20 | 1.44 | 1.8 | " | |
| East Mediterranean | 5–200 | summer 07 | 18.41 | 38.79 | 1.53 | 0.50 | 4.56 | 3.06 | 0.33 | [49] | |
| 500–1000 | " | 14.01 | 38.81 | 0.66 | 0.52 | 0.33 | 1.27 | 2.01 | " | ||
| 1500– 2900 | " | 13.95 | 38.75 | 1.22 | 0.58 | 0.78 | 2.10 | 1.56 | " | ||
| West Mediterranean | 5–200 | 2.18 | 0.88 | 0.51 | [59] | ||||||
| 500–1000 | 1.04 | 0.51 | 0.35 | " | |||||||
| West Mediterranean | 1500–3500 | 0.66 | 0.41 | 0.31 | " | ||||||
| East Mediterranean | 5–200 | 0.08 | 0.31 | 0.20 | [59] | ||||||
| 500–1000 | 0.09 | 0.10 | 0.13 | " | |||||||
| 1500– 2900 | 0.05 | 0.13 | 0.03 | " | |||||||
| North Adriatic Sea | gen-60 | coastal station | seasonal study | 2.2–1701 | 0.3–114 | 0.8 -1770 | [70] | ||||
| North Adriatic Sea | 1–60 | coastal station | spring-summer | 115– 67.5 | 2.1–5.2 | 13.6 | 22.14 | 4.96 | [73] | ||
| North Adriatic Sea | autumn | 2.04–4.64 | [38] | ||||||||
| North Adriatic Sea | 1–10 | coastal station | spring-autumn | 70–4640 | [37] | ||||||
| South Adriatic Sea | 5–100 | convection site | seasonal study | 13.8–15.6 | 38.71–38.74 | 0.01–7.86 | 0.003–3.5 | 0.05–31.96 | [57] | ||
| 100–800 | " | 13.6–13.6 | 38.74–38.77 | 0.01–2.66 | 0.003–0.98 | 0.003–6.47 | " | ||||
| " | 800–1200 | " | 12.9–13.1 | 38.72–38.73 | 0.01–0.15 | 0.003–0.95 | 0.08–17.96 | " | |||
| South Adriatic Sea | 0–30 | Gulf of Manfredonia | spring 03 | 18.44 | 37.92 | 13.99 | 1.13 | 43.32 | 12.4 | 0.32 | [58] |
| 0–30 | offshore waters | " | 17.25 | 38.73 | 31.18 | 0.48 | 43,28 | 72.51 | 0.72 | " | |
| >30 | " | " | 14.11 | 38.70 | 19.45 | 0.26 | 23.49 | 75.97 | 0.83 | " | |
| North Atlantic | surface | 4.2–24.9 | [27] | ||||||||
| South Atlantic | surface | 11.5–151.4 | " | ||||||||
| Atlantic ocean | 100–4500 | 0.2–13.8 | 0.05–0.25 | 0.14–1.3 | [34] | ||||||
| subtropical Atlantic | 100–4500 | 0.6–9.2 | 0.02–0.07 | 0.04–3.7 | [67] |
