Citation: Amir Hedayati Aghmashhadi, Giuseppe T. Cirella, Samaneh Zahedi, Azadeh Kazemi. Water resource policy support system of the Caspian Basin[J]. AIMS Environmental Science, 2019, 6(4): 242-261. doi: 10.3934/environsci.2019.4.242

| [1] | Mitchell M, Curtis A, Sharp E, et al. (2012) Directions for social research to underpin improved groundwater management. J Hydrol 448: 223–231. |
| [2] |
Conrad SA, Yates D (2018) Coupling stated preferences with a hydrological water resource model to inform water policies for residential areas in the Okanagan Basin, Canada. J Hydrol 564: 846–858. doi: 10.1016/j.jhydrol.2018.07.031

|
| [3] |
Hanjra MA, Qureshi ME (2010) Global water crisis and future food security in an era of climate change. Food Policy 35: 365–377. doi: 10.1016/j.foodpol.2010.05.006
|
| [4] |
Gu S, Jenkins A, Gao SJ, et al. (2017) Ensuring water resource security in China: The need for advances in evidence-based policy to support sustainable management. Environ Sci Policy 75: 65–69. doi: 10.1016/j.envsci.2017.05.008
|
| [5] |
Freund A, Aydin NY, Zeckzer D, et al. (2017) A Decision-Support System for Sustainable Water Distribution System Planning. IEEE Comput Graph Appl 37: 44–55. doi: 10.1109/MCG.2015.120
|
| [6] |
Gastélum JR, Valdés JB, Stewart S (2009) A Decision Support System to Improve Water Resources Management in the Conchos Basin. Water Resour Manag 23: 1519–1548. doi: 10.1007/s11269-008-9339-4
|
| [7] | Wilder M (2010) Water governance in Mexico: Political and economic apertures and a shifting state-citizen relationship. Ecol Soc 15: 1–18. |
| [8] |
Rowley HV, Peters GM, Lundie S, et al. (2012) Aggregating sustainability indicators: Beyond the weighted sum. J Environ Manage 111: 24–33. doi: 10.1016/j.jenvman.2012.05.004
|
| [9] | Hedayati Aghmashhadi A (2018) Water Resource Policy Making based on the Pressures Management: Policy, Planning and Management of Water Resource in the Arid and Semi-Arid Regions. Beau-Bassin, Mauritius: LAP Publishing. |
| [10] |
Yu Y, Chen X, Huttner P, et al. (2018) Model based decision support system for land use changes and socio-economic assessments. J Arid Land 10: 169–182. doi: 10.1007/s40333-018-0091-1
|
| [11] |
Loucks DP, Kindler J, Fedra K (1985) Interactive Water Resources Modeling and Model Use: An Overview. Water Resour Res 21: 95–102. doi: 10.1029/WR021i002p00095
|
| [12] | Georgakakos AP (2007) Decision support systems for integrated water resources management with an application to the Nile Basin. In: Castelletti A, Soncini-Sessa R, Topics on System Analysis and Integrated Water Resources Management. Amsterdam: Elsevier Science, 99–116. |
| [13] | Serrat-Capdevila A, Valdes BJ, Gupta HV (2011) Decision Support Systems in Water Resources Planning and Management: Stakeholder Participation and the Sustainable Path to Science-Based Decision Making. In: Jao C, Efficient Decision Support Systems: Practice and Challenges From Current to Future. Rijeka, Croatia: InTech. |
| [14] | Othman F, Naseri M (2008) Decision Support Systems in Water Resources Management. In: Proceedings of the 9th Asia Pasific Industrial Engineering Management Systems Conference. Bali, Indonesia : APIEMS 2008, 1772–1780. |
| [15] | Neuman J (2010) Are We There Yet? Weary Travelers on the Long Road to Water Policy Reform. Nat Resour J 211: 139–166. |
| [16] |
Gutiãrrez RA (2010) When Experts Do Politics: Introducing Water Policy Reform in Brazil. Governance 23: 59–88. doi: 10.1111/j.1468-0491.2009.01467.x
|
| [17] |
Lein H, Tagseth M (2009) Tanzanian water policy reforms-between principles and practical applications. Water Policy 11: 203–220. doi: 10.2166/wp.2009.024
|
| [18] |
Nhapi I (2009) The water situation in Harare, Zimbabwe: A policy and management problem. Water Policy 11: 221–235. doi: 10.2166/wp.2009.018
|
| [19] |
Biswas AK, Tortajada C (2010) Future Water Governance: Problems and Perspectives. Int J Water Resour Dev 26: 129–139. doi: 10.1080/07900627.2010.488853
|
| [20] |
Schlueter M, Hirsch D, Pahl-Wostl C (2010) Coping with change: Responses of the Uzbek water management regime to socio-economic transition and global change. Environ Sci Policy 13: 620–636. doi: 10.1016/j.envsci.2010.09.001
|
| [21] |
Akter S, Grafton RQ, Merritt WS (2014) Integrated hydro-ecological and economic modeling of environmental flows: Macquarie Marshes, Australia. Agric Water Manag 145: 98–109. doi: 10.1016/j.agwat.2013.12.005
|
| [22] |
Hurd BH, Coonrod J (2012) Hydro-economic consequences of climate change in the upper Rio Grande. Clim Res 53: 103–118. doi: 10.3354/cr01092
|
| [23] | Koundouri P, Reppas D, Skianis V (2015) Socio-Economics and Water Management: Revisiting the Contribution of Economics in the Implementation of the Water Framework Directive in Greece. Berlin: Springer, 357–377. |
| [24] |
Lanini S, Courtois N, Giraud F, et al. (2004) Socio-hydrosystem modelling for integrated water-resources management-the Hérault catchment case study, southern France. Environ Model Softw 19: 1011–1019. doi: 10.1016/j.envsoft.2003.11.004
|
| [25] |
Słowiński R (1986) A multicriteria fuzzy linear programming method for water supply system development planning. Fuzzy Sets Syst 19: 217–237. doi: 10.1016/0165-0114(86)90052-7
|
| [26] |
Huang GH (1998) A hybrid inexact-stochastic water management model. Eur J Oper Res 107: 137–158. doi: 10.1016/S0377-2217(97)00144-6
|
| [27] |
Jairaj PG, Vedula S (2000) Multireservoir System Optimization using Fuzzy Mathematical Programming. Water Resour Manag 14: 457–472. doi: 10.1023/A:1011117918943
|
| [28] |
Luo B (2003) Adaption to Climate Change through Water Trading under Uncertainty-An Inexact Two-Stage Nonlinear Programming Approach. J Environ Informatics 2: 58–68. doi: 10.3808/jei.200300022
|
| [29] |
Maqsood I, Huang GH, Yeomans JS (2005) An interval-parameter fuzzy two-stage stochastic program for water resources management under uncertainty. Eur J Oper Res 167: 208–225. doi: 10.1016/j.ejor.2003.08.068
|
| [30] |
Li YP, Huang GH, Nie SL, et al. (2007) ITCLP: An inexact two-stage chance-constrained program for planning waste management systems. Resour Conserv Recycl 49: 284–307. doi: 10.1016/j.resconrec.2006.03.017
|
| [31] | Wu SM, Huang GH, Guo HC (1997) An interactive inexact-fuzzy approach for multiobjective planning of water resource systems. Water Sci Technol 36: 235–242. |
| [32] |
Seifi A, Hipel KW (2001) Interior-Point Method for Reservoir Operation with Stochastic Inflows. J Water Resour Plan Manag 127: 48–57. doi: 10.1061/(ASCE)0733-9496(2001)127:1(48)
|
| [33] |
Wang S, Huang GH, Lu HW, et al. (2011) An interval-valued fuzzy linear programming with infinite α-cuts method for environmental management under uncertainty. Stoch Environ Res Risk Assess 25: 211–222. doi: 10.1007/s00477-010-0432-x
|
| [34] |
Huang GH (1996) IPWM: An Interval Parameter Water Quality Management Model. Eng Optim 26: 79–103. doi: 10.1080/03052159608941111
|
| [35] |
Bender MJ, Simonovic SP (2000) A fuzzy compromise approach to water resource systems planning under uncertainty. Fuzzy Sets Syst 115: 35–44. doi: 10.1016/S0165-0114(99)00025-1
|
| [36] |
Simonovic SP, Fahmy H (1999) A new modeling approach for water resources policy analysis. Water Resour Res 35: 295–304. doi: 10.1029/1998WR900023
|
| [37] | Faye RM, Sawadogo S, Mora-Camino F (2005) Flexible management of water resource systems. Appl Math Comput 167: 516–527. |
| [38] |
Lee CS, Chang SP (2005) Interactive fuzzy optimization for an economic and environmental balance in a river system. Water Res 39: 221–231. doi: 10.1016/j.watres.2004.09.013
|
| [39] |
Edirisinghe NCP, Patterson EI, Saadouli N (2000) Capacity Planning Model for a Multipurpose Water Reservoir with Target-Priority Operation. Ann Oper Res 100: 273–303. doi: 10.1023/A:1019200623139
|
| [40] |
Azaiez MN (2002) A model for conjunctive use of ground and surface water with opportunity costs. Eur J Oper Res 143: 611–624. doi: 10.1016/S0377-2217(01)00339-3
|
| [41] |
Pallottino S, Sechi GM, Zuddas P (2005) A DSS for water resources management under uncertainty by scenario analysis. Environ Model Softw 20: 1031–1042. doi: 10.1016/j.envsoft.2004.09.012
|
| [42] |
Li YP, Huang GH, Nie SL (2006) An interval-parameter multi-stage stochastic programming model for water resources management under uncertainty. Adv Water Resour 29: 776–789. doi: 10.1016/j.advwatres.2005.07.008
|
| [43] |
Nasiri F, Maqsood I, Huang G, et al. (2007) Water Quality Index: A Fuzzy River-Pollution Decision Support Expert System. J Water Resour Plan Manag 133: 95–105. doi: 10.1061/(ASCE)0733-9496(2007)133:2(95)
|
| [44] |
Carey M, Baraer M, Mark BG, et al. (2014) Toward hydro-social modeling: Merging human variables and the social sciences with climate-glacier runoff models (Santa River, Peru). J Hydrol 518: 60–70. doi: 10.1016/j.jhydrol.2013.11.006
|
| [45] |
Giupponi C, Sgobbi A (2013) Decision Support Systems for Water Resources Management in Developing Countries: Learning from Experiences in Africa. Water 5: 798–818. doi: 10.3390/w5020798
|
| [46] |
Soncini-Sessa R, Castelletti A, Weber E (2003) A DSS for planning and managing water reservoir systems. Environ Model Softw 18: 395–404. doi: 10.1016/S1364-8152(03)00035-5
|
| [47] |
Li YP, Huang GH, Huang YF, et al. (2009) A multistage fuzzy-stochastic programming model for supporting sustainable water-resources allocation and management. Environ Model Softw 24: 786–797. doi: 10.1016/j.envsoft.2008.11.008
|
| [48] |
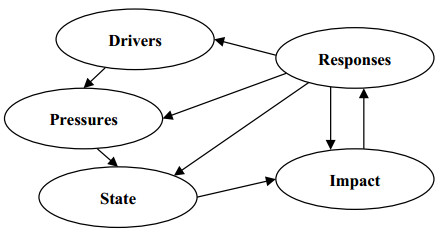
Spangenberg JH, Douguet J-M, Settele J, et al. (2015) Escaping the lock-in of continuous insecticide spraying in rice: Developing an integrated ecological and socio-political DPSIR analysis. Ecol Modell 295: 188–195. doi: 10.1016/j.ecolmodel.2014.05.010
|
| [49] | Smeets E, Weterings R (1999) Environmental indicators: Typology and overview. Copenhagen: European Environment Agency Press. |
| [50] | Gabrielsen P, Bosch P (2003) Environmental indicators: Typology and overview. Copenhagen: European Environment Agency Intern. Work. Pap. |
| [51] | European Environment Agency (2006) EEA Glossary. Brussels: European Commission. |
| [52] | Voogd H (1983) Multicriteria evaluation for urban and regional planning. Delft: Delftsche Uitgevers Maatschappij B.V. |
| [53] |
Carver SJ (1991) Integrating multi-criteria evaluation with geographical information systems. Int J Geogr Inf Syst 5: 321–339. doi: 10.1080/02693799108927858
|
| [54] | Eastman JR (2012) IDRISI Selva manual. Worcester, MA, USA: Clark University Press. |
| [55] | Malczewski J (1999) GIS and multicriteria decision analysis. New York: John Wiley and Sons, Inc. |
| [56] | Shahabi H, Niyazi C (2009) Effecting factors in relief and rescue stations site selection by using weighted linear combination, case study: Saghez-Sanandaj road. In: Proceedings of the Eighth International Conference on Geomatics. Tehran: ICG 2009. |
| [57] | Burrough PA (1990) Methods of Spatial Analysis in GIS. Int J Geogr Inform Syst 4: 221–223. |
| [58] | Saaty TL (2001) Fundamentals of the Analytic Hierarchy Process. Dordrecht: Springer, 15–35. |
| [59] |
Deng H (1999) Multicriteria analysis with fuzzy pairwise comparison. Int J Approx Reason 21: 215–231. doi: 10.1016/S0888-613X(99)00025-0
|
| [60] |
Boroushaki S, Malczewski J (2008) Implementing an extension of the analytical hierarchy process using ordered weighted averaging operators with fuzzy quantifiers in ArcGIS. Comput Geosci 34: 399–410. doi: 10.1016/j.cageo.2007.04.003
|
| [61] |
Linkov I, Satterstrom FK, Steevens J, et al. (2007) Multi-criteria decision analysis and environmental risk assessment for nanomaterials. J Nanoparticle Res 9: 543–554. doi: 10.1007/s11051-007-9211-0
|
| [62] | Alam Tabriz A, Bagherzade Azar M (2009) Combining AHP and TOPSIS Adjusted for supplier selection strategy. Res Manag 6: 149–181. |
| [63] | Ghodsi Poor H (2010) Analytical Hierarchy Process (AHP). Tehran: Amirkabir University of Technology Press. |
| [64] | Razmi J, Sadegh Amal Nik M, Hashemi M (2008) Supplier selection techniques using fuzzy analytic network process. Tehran Uni Eng 42: 935–946. |
| [65] |
Leung LC, Cao D (2000) On consistency and ranking of alternatives in fuzzy AHP. Eur J Oper Res 124: 102–113. doi: 10.1016/S0377-2217(99)00118-6
|
| [66] | Hansen HS (2005) GIS-based Multi-Criteria Analysis of Wind Farm Development. In: Hauska H, Tveite H, ScanGis 2005: Proceedings of the Tenth Scandinavian Research Conference on Geographical Information Science. Stockholm: Department of Planning and Environment, 75–87. |
| [67] |
Lee S (2007) Application and verification of fuzzy algebraic operators to landslide susceptibility mapping. Environ Geol 52: 615–623. doi: 10.1007/s00254-006-0491-y
|
| [68] | Kabir S, Edifor E, Walker M, et al. (2014) Quantification of Temporal Fault Trees Based on Fuzzy Set Theory. Springer, Cham, pp 255–264 |
| [69] | Ghosh JK, Bhattacharya D, Sharma SK (2012) Fuzzy Knowledge Based GIS for Zonation of Landslide Susceptibility. In: Zamojski W, Mazurkiewicz J, Sugier J, et al., Proceedings of the Ninth International Conference on Dependability and Complex Systems DepCoS-RELCOMEX. Brunów, Poland: Springer, Cham. |
| [70] |
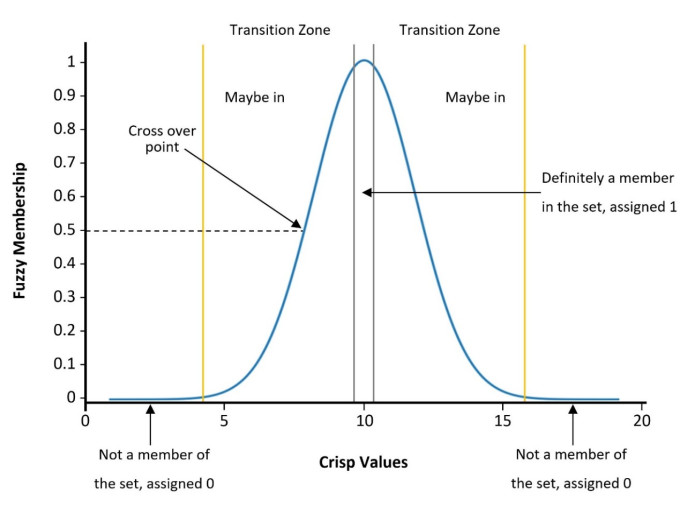
Dombi J (1990) Membership function as an evaluation. Fuzzy Sets Syst 35: 1–21. doi: 10.1016/0165-0114(90)90014-W
|
| [71] | Keller CP (1995) Geographic information systems for geoscientists: Modelling with GIS. Iny Graeme F. Bonham-Carter, Pergamon, Computer Methods in the Geosciences, Volume 13, 1994, 398 p., incl. bibliographic references and index, US $40.00. Comput Geosci 21: 1110–1112. |
| [72] |
Zimmermann HJ, Zysno P (1980) Latent connectives in human decision making. Fuzzy Sets Syst 4: 37–51. doi: 10.1016/0165-0114(80)90062-7
|
| [73] |
Atkinson DM, Deadman P, Dudycha D, et al. (2005) Multi-criteria evaluation and least cost path analysis for an arctic all-weather road. Appl Geogr 25: 287–307. doi: 10.1016/j.apgeog.2005.08.001
|
| [74] | Salari M, Bagherpour M, Wang J (2014) A novel earned value management model using Z-number. Int J Appl Decis Sci 7: 97–119. |
| [75] |
Önüt S, Kara SS, Işik E (2009) Long term supplier selection using a combined fuzzy MCDM approach: A case study for a telecommunication company. Expert Syst Appl 36: 3887–3895. doi: 10.1016/j.eswa.2008.02.045
|
| [76] |
Gogus O, Boucher TO (1998) Strong transitivity, rationality and weak monotonicity in fuzzy pairwise comparisons. Fuzzy Sets Syst 94: 133–144. doi: 10.1016/S0165-0114(96)00184-4
|
| [77] | Lin H, Kao J, Li K, et al. (1996) Fuzzy GIS assisted landfill siting analysis. In: Proceedings of the Twelfth International Conference on Solid Waste Technology and Management. Philadelphia, PA, USA: Journal of Solid Waste Technology and Management, 14–17. |
| [78] | Valizadeh K, Shababi H (2009) Necessities of GIS usage in urban water management at the time of Natural accidents (Case Study: Saqqez City). In: Proceeding of the Fourth International Conference on Geographic Information Systems. Paris: ICGIS. |
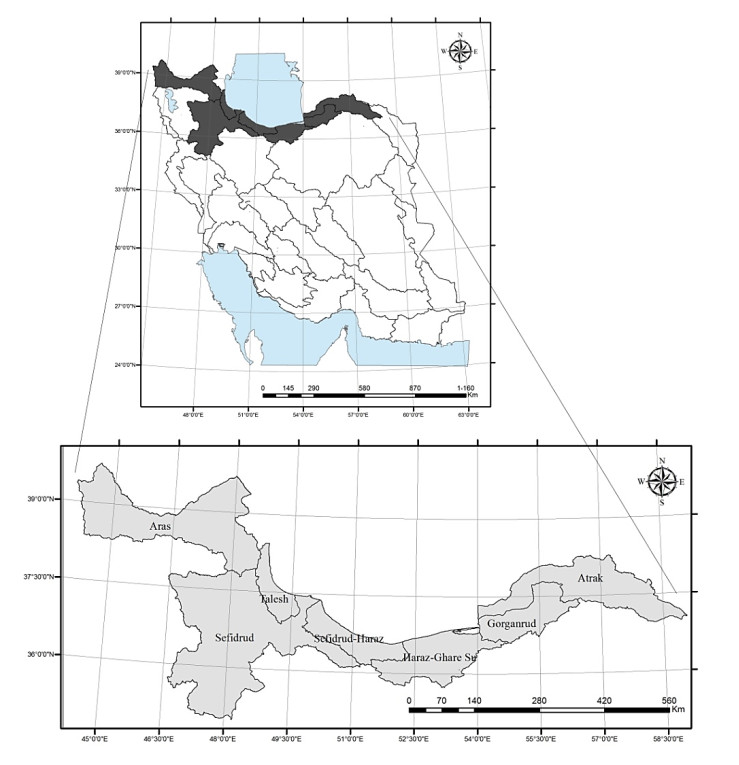
| [79] | Ministry of Energy (2013) Water Master Plan Udate in the Aras, Urmia, Talesh, Sefidrud, Sefidrud-Haraz, Haraz-Ghareh Su, Gorganrud-Ghareh Su, Atrak. Ministry of Energy, Water and Abfa Deputy. Tehran: Office of Planning and Abfa Macro Water. |
| [80] | Statistical Center of Iran (2012) Iran Census of Population and Housing, 2011. Tehran: SCI Press. |
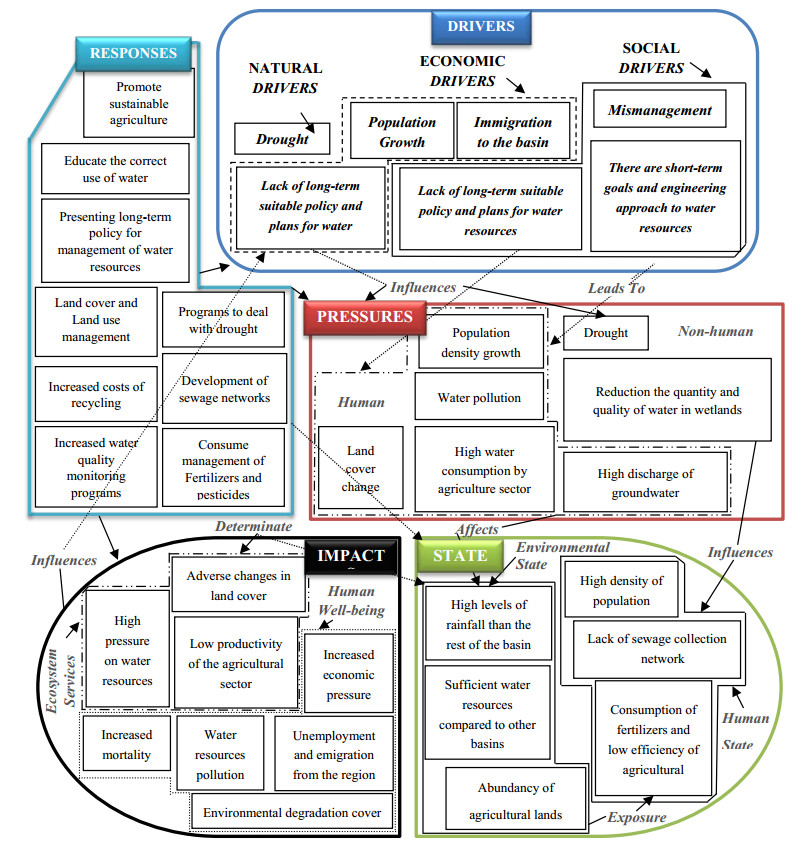
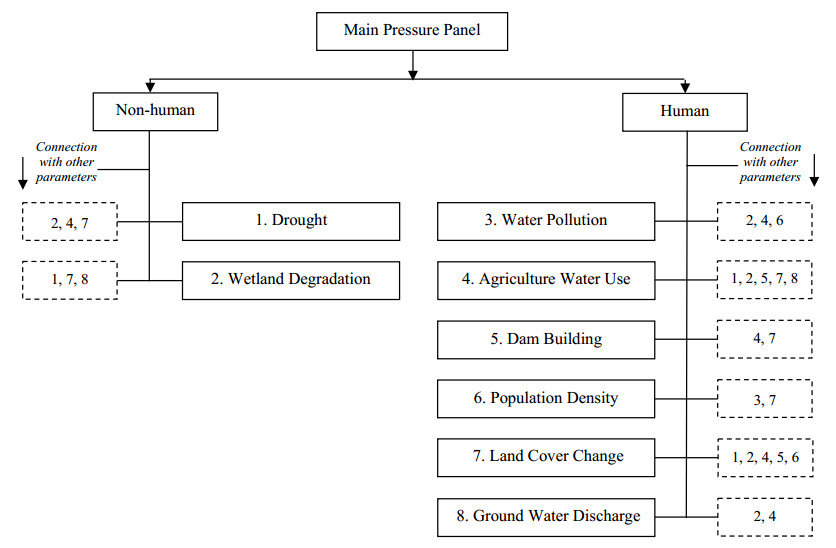
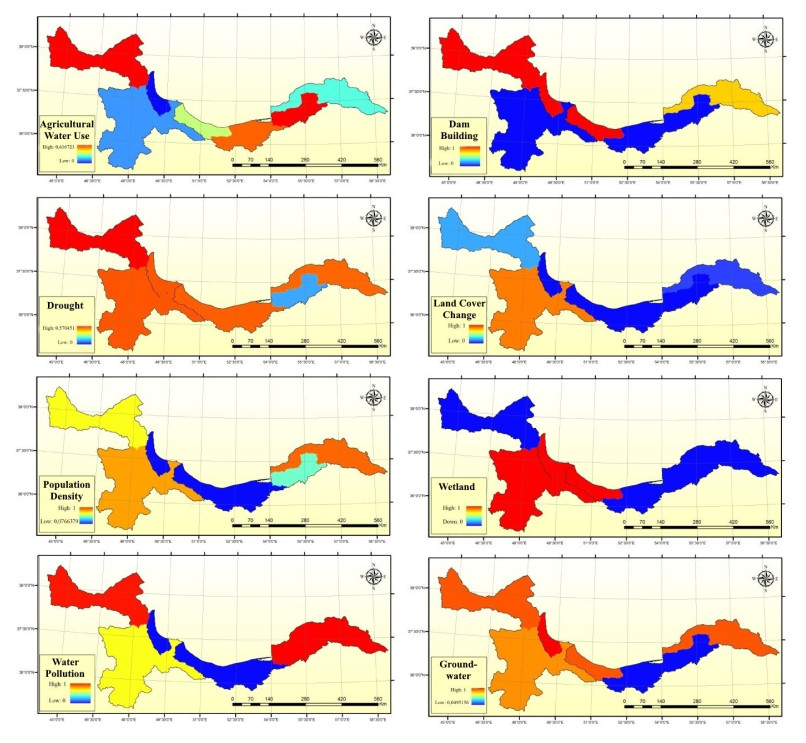
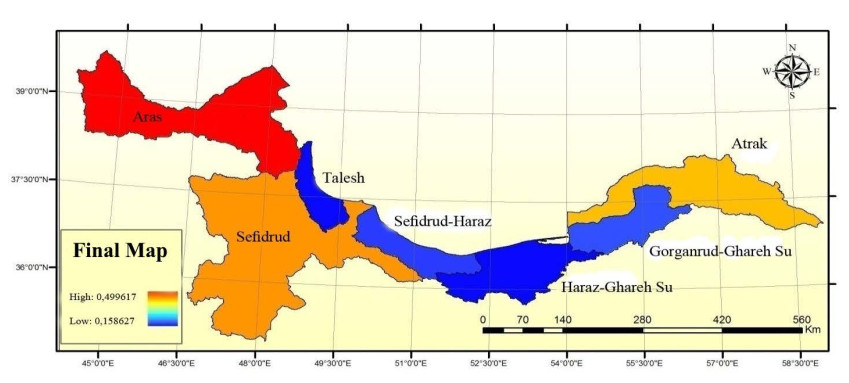
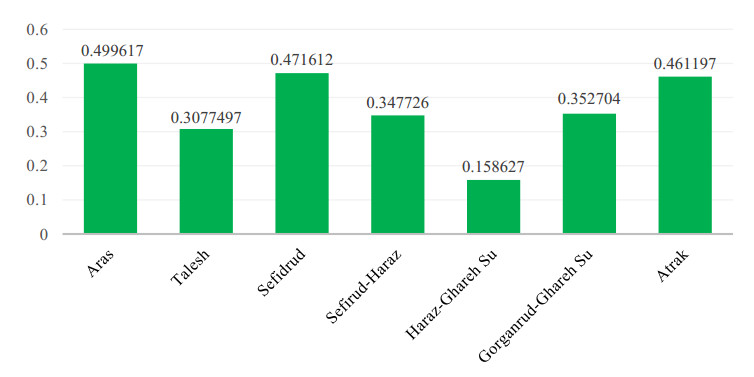
Figures(9) / Tables(5)

Amir Hedayati Aghmashhadi, Giuseppe T. Cirella, Samaneh Zahedi, Azadeh Kazemi. Water resource policy support system of the Caspian Basin[J]. AIMS Environmental Science, 2019, 6(4): 242-261. doi: 10.3934/environsci.2019.4.242







 DownLoad:
DownLoad: