Citation: Saad S. Alrwashdeh, Falah M. Alsaraireh, Mohammad A. Saraireh, Henning Markötter, Nikolay Kardjilov, Merle Klages, Joachim Scholta, Ingo Manke. In-situ investigation of water distribution in polymer electrolyte membrane fuel cells using high-resolution neutron tomography with 6.5 µm pixel size[J]. AIMS Energy, 2018, 6(4): 607-614. doi: 10.3934/energy.2018.4.607

| [1] | Hoogers G (2003) Fuel Cell Technology Handbook. Boca Raton, FL: CRC Press LLC. |
| [2] | Vielstich W, Lamm A, Gasteiger HA (2003) Handbook of fuel cells–fundamentals, technology and applications. Chichester: John Wiley & Sons. |
| [3] |
Alrwashdeh SS, Manke I, Markötter H, et al. (2017) In operando quantification of three-dimensional water distribution in nanoporous carbon-based layers in polymer electrolyte membrane fuel cells. ACS Nano 11: 5944–5949. doi: 10.1021/acsnano.7b01720

|
| [4] |
Alrwashdeh SS, Manke I, Markötter H, et al. (2017) Neutron radiographic in operando investigation of water transport in polymer electrolyte membrane fuel cells with channel barriers. Energ Convers Manage 148: 604–610. doi: 10.1016/j.enconman.2017.06.032
|
| [5] |
Alrwashdeh SS, Manke I, Markötter H, et al. (2017) Improved performance of polymer electrolyte membrane fuel cells with modified microporous layer structures. Energy Technol 5: 1612–1618. doi: 10.1002/ente.201700005
|
| [6] |
Saad SA, Henning M, Haußmann J, et al. (2017) Investigation of water transport in newly developed micro porous layers for polymer electrolyte membrane fuel cells. Appl Microscopy 47: 101–104. doi: 10.9729/AM.2017.47.3.101
|
| [7] |
Mehta V, Cooper JS (2003) Review and analysis of PEM fuel cell design and manufacturing. J Power Sources 114: 32–53. doi: 10.1016/S0378-7753(02)00542-6
|
| [8] | Carrette L, Friedrich KA, Stimming U (2000) Fuel cells: principles, types, fuels, and applications: WILEY-VCH Verlag GmbH, Weinheim. |
| [9] |
Barelli L, Bidini G, Gallorini F, et al. (2012) Dynamic analysis of PEMFC-based CHP systems for domestic application. Appl Energ 91: 13–28. doi: 10.1016/j.apenergy.2011.09.008
|
| [10] |
Gigliucci G, Petruzzi L, Cerelli E, et al. (2004) Demonstration of a residential CHP system based on PEM fuel cells. J Power Sources 131: 62–68. doi: 10.1016/j.jpowsour.2004.01.010
|
| [11] |
Krüger P, Markötter H, Haußmann J, et al. (2011) Synchrotron X-ray tomography for investigations of water distribution in polymer electrolyte membrane fuel cells. J Power Sources 196: 5250–5255. doi: 10.1016/j.jpowsour.2010.09.042
|
| [12] |
Manke I, Hartnig C, Kardjilov N, et al. (2009) In-situ investigation of the water distribution in PEM fuel cells by neutron radiography and tomography. Mater Test 51: 219–226. doi: 10.3139/120.110015
|
| [13] |
Alrwashdeh SS, Markötter H, Haußmann J, et al. (2016) Investigation of water transport dynamics in polymer electrolyte membrane fuel cells based on high porous micro porous layers. Energy 102: 161–165. doi: 10.1016/j.energy.2016.02.075
|
| [14] |
Cindrella L, Kannan AM (2009) Membrane electrode assembly with doped polyaniline interlayer for proton exchange membrane fuel cells under low relative humidity conditions. J Power Sources 193: 447–453. doi: 10.1016/j.jpowsour.2009.04.002
|
| [15] |
Cindrella L, Kannan AM, Ahmad R, et al. (2009) Surface modification of gas diffusion layers by inorganic nanomaterials for performance enhancement of proton exchange membrane fuel cells at low RH conditions. Int J Hydrogen Energ 34: 6377–6383. doi: 10.1016/j.ijhydene.2009.05.086
|
| [16] |
Mohanraju K, Sreejith V, Ananth R, et al. (2015) Enhanced electrocatalytic activity of PANI and CoFe2O4/PANI composite supported on graphene for fuel cell applications. J Power Sources 284: 383–391. doi: 10.1016/j.jpowsour.2015.03.025
|
| [17] |
Cindrella L, Kannan AM, Lin JF, et al. (2009) Gas diffusion layer for proton exchange membrane fuel cells-A review. J Power Sources 194: 146–160. doi: 10.1016/j.jpowsour.2009.04.005
|
| [18] |
Chevalier S, Ge N, George MG, et al. (2017) Synchrotron X-ray radiography as a highly precise and accurate method for measuring the spatial distribution of liquid water in operating polymer electrolyte membrane fuel cells. J Electrochem Soc 164: F107–F114. doi: 10.1149/2.0041702jes
|
| [19] |
Ge N, Chevalier S, Lee J, et al. (2017) Non-isothermal two-phase transport in a polymer electrolyte membrane fuel cell with crack-free microporous layers. Int J Heat Mass Transfer 107: 418–431. doi: 10.1016/j.ijheatmasstransfer.2016.11.045
|
| [20] |
Antonacci P, Chevalier S, Lee J, et al. (2015) Feasibility of combining electrochemical impedance spectroscopy and synchrotron X-ray radiography for determining the influence of liquid water on polymer electrolyte membrane fuel cell performance. Int J Hydrogen Energ 40: 16494–1502. doi: 10.1016/j.ijhydene.2015.10.008
|
| [21] |
Lee J, Hinebaugh J, Bazylak A (2013) Synchrotron X-ray radiographic investigations of liquid water transport behavior in a PEMFC with MPL-coated GDLs. J Power Sources 227: 123–130. doi: 10.1016/j.jpowsour.2012.11.006
|
| [22] |
Arlt T, Grothausmann R, Manke I, et al. (2013) Tomographic methods for fuel cell research. Mater Test 55: 207–213. doi: 10.3139/120.110429
|
| [23] |
Eberhardt SH, Marone F, Stampanoni M, et al. (2016) Operando X-ray tomographic microscopy imaging of HT-PEFC: A comparative study of phosphoric acid electrolyte migration. J Electrochem Soc 163: F842–F847. doi: 10.1149/2.0801608jes
|
| [24] |
Arlt T, Klages M, Messerschmidt M, et al. (2017) Influence of artificially aged gas diffusion layers on the water management of polymer electrolyte membrane fuel cells analyzed with in-operando synchrotron imaging. Energy 118: 502–511. doi: 10.1016/j.energy.2016.10.061
|
| [25] | Chevalier S, Ge N, Lee J, et al. (2017) Novel electrospun gas diffusion layers for polymer electrolyte membrane fuel cells: Part II. In operando synchrotron imaging for microscale liquid water transport characterization. J Power Sources 352: 281–290. |
| [26] |
Matsui H, Ishiguro N, Uruga T, et al. (2017) Operando 3D visualization of migration and degradation of a platinum cathode catalyst in a polymer electrolyte fuel cell. Angew Chem Int Ed Engl 56: 9371–9375. doi: 10.1002/anie.201703940
|
| [27] |
Kardjilov N, Hilger A, Manke I, et al. (2011) Neutron tomography instrument CONRAD at HZB. Nucl Instrum Meth A 651: 47–52. doi: 10.1016/j.nima.2011.01.067
|
| [28] |
Kardjilov N, Manke I, Hilger A, et al. (2011) Neutron imaging in materials science. Mater Today 14: 248–256. doi: 10.1016/S1369-7021(11)70139-0
|
| [29] | Kardjilov N, Hilger A, Manke I, et al. (2015) Imaging with cold neutrons at the CONRAD-2 Facility. In: Lehmann EH, Kaestner AP, Mannes D, editors, Proceedings of the 10th World Conference on Neutron Radiography. Amsterdam: Elsevier Science Bv, 60–66. |
| [30] |
Kardjilov N, Hilger A, Manke I, et al. (2016) CONRAD-2: the new neutron imaging instrument at the Helmholtz-Zentrum Berlin. J Appl Crystallogr 49: 195–202. doi: 10.1107/S1600576715023353
|
| [31] | Williams SH, Hilger A, Kardjilov N, et al. (2012) Detection system for microimaging with neutrons. J Instrum 7: 1–25. |
| [32] |
Kardjilov N, Dawson M, Hilger A, et al. (2011) A highly adaptive detector system for high resolution neutron imaging. Nucl Instrum Meth A 651: 95–99. doi: 10.1016/j.nima.2011.02.084
|
| [33] |
Totzke C, Manke I, Hilger A, et al. (2011) Large area high resolution neutron imaging detector for fuel cell research. J Power Sources 196: 4631–4637. doi: 10.1016/j.jpowsour.2011.01.049
|
| [34] | Banhart J (2008) Advanced tomographic methods in materials research and engineering. Oxford, UK: Oxford University, Press. |
| [35] | Manke I, Hartnig C, Kardjilov N, et al. (2008) Characterization of water exchange and two-phase flow in porous gas diffusion materials by hydrogen-deuterium contrast neutron radiography. Appl Phys Lett 92: 337–347. |
| [36] |
Cho KT, Mench MM (2012) Investigation of the role of the micro-porous layer in polymer electrolyte fuel cells with hydrogen deuterium contrast neutron radiography. Phys Chem Chem Phys 14: 4296–4302. doi: 10.1039/c2cp23686a
|
| [37] |
Haussmann J, Markotter H, Alink R, et al. (2013) Synchrotron radiography and tomography of water transport in perforated gas diffusion media. J Power Sources 239: 611–622. doi: 10.1016/j.jpowsour.2013.02.014
|
| [38] |
Weber AZ, Borup RL, Darling RM, et al. (2014) A critical review of modeling transport phenomena in polymer-electrolyte fuel cells. J Electrochem Soc 161: F1254–F1299. doi: 10.1149/2.0751412jes
|
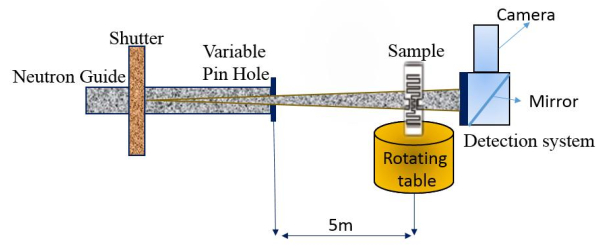

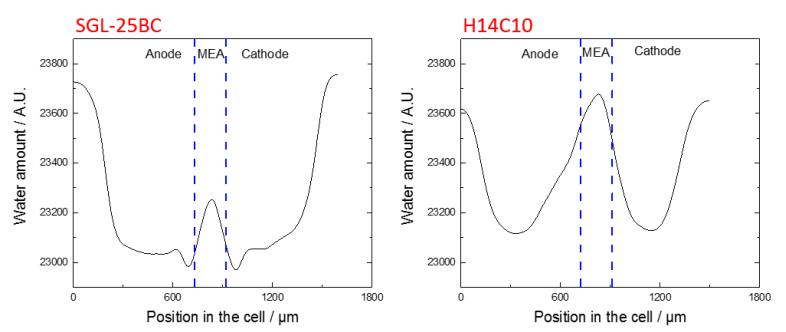
Figures(3)

Saad S. Alrwashdeh, Falah M. Alsaraireh, Mohammad A. Saraireh, Henning Markötter, Nikolay Kardjilov, Merle Klages, Joachim Scholta, Ingo Manke. In-situ investigation of water distribution in polymer electrolyte membrane fuel cells using high-resolution neutron tomography with 6.5 µm pixel size[J]. AIMS Energy, 2018, 6(4): 607-614. doi: 10.3934/energy.2018.4.607







 DownLoad:
DownLoad: