Citation: Albert W. Pilkington IV, Justin Legleiter. Challenges in understanding the structure/activity relationship of Aβ oligomers[J]. AIMS Biophysics, 2019, 6(1): 1-22. doi: 10.3934/biophy.2019.1.1

| [1] |
Knowles TP, Vendruscolo M, Dobson CM (2014) The amyloid state and its association with protein misfolding diseases. Nat Rev Mol Cell Biol 15: 384–396. doi: 10.1038/nrm3810

|
| [2] |
Hardy JA, Higgins GA (1992) Alzheimers disease-the amyloid cascade hyopothesis. Science 256: 184–185. doi: 10.1126/science.1566067
|
| [3] |
Arriagada PV, Growdon JH, Hedleywhyte ET, et al. (1992) Neurofibrillary tangles but not senile plaques parellel duration and severity of Alzheimers disease. Neurology 42: 631–639. doi: 10.1212/WNL.42.3.631
|
| [4] |
Terry RD, Masliah E, Salmon DP, et al. (1991) Physical basis of cognitve alterations in Alzheimers disease-synapse loss is the major correlate of cognitive impairment. Ann Neurol 30: 572–580. doi: 10.1002/ana.410300410
|
| [5] |
Viola KL, Sbarboro J, Sureka R, et al. (2015) Towards non-invasive diagnostic imaging of early-stage Alzheimer's disease. Nat Nanotechnol 10: 91–98. doi: 10.1038/nnano.2014.254
|
| [6] | Hyman B, Tanzi R (1992) Amyloid, dementia and Alzheimer's disease. Curr Opin Neurol Neurosur 5: 88–93. |
| [7] |
Cummings BJ, Pike CJ, Shankle R, et al. (1996) b-Amyloid deposition and other measures of neuropathology predict cognitive status in Alzheimer's disease. Neurobiol Aging 17: 921–933. doi: 10.1016/S0197-4580(96)00170-4
|
| [8] |
Cummings JL, Morstorf T, Zhong K (2014) Alzheimer's disease drug-development pipeline: Few candidates, frequent failures. Alzheimers Res Ther 6: 37. doi: 10.1186/alzrt269
|
| [9] |
Goure WF, Krafft GA, Jerecic J, et al. (2014) Targeting the proper amyloid-β neuronal toxins: A path forward for Alzheimer's disease immunotherapeutics. Alzheimers Res Ther 6: 42. doi: 10.1186/alzrt272
|
| [10] |
Karran E, Hardy J (2014) A critique of the drug discovery and phase 3 clinical programs targeting the amyloid hypothesis for Alzheimer disease. Ann Neurol 76: 185–205. doi: 10.1002/ana.24188
|
| [11] |
Karran E, Hardy J (2014) Antiamyloid therapy for Alzheimer's disease-are we on the right road? New Engl J Med 370: 377–378. doi: 10.1056/NEJMe1313943
|
| [12] |
Frackowiak J, Zoltowska A, Wisniewski HM (1994) Nonfibrillar β-amyloid protein is associated with smooth-muscle cells of vessel walls in Alzheimer disease. J Neuropath Exp Neur 53: 637–645. doi: 10.1097/00005072-199411000-00011
|
| [13] |
Oda T, Pasinetti GM, Osterburg HH, et al. (1994) Purification and characterization of brain clusterin. Biochem Bioph Res Co 204: 1131–1136. doi: 10.1006/bbrc.1994.2580
|
| [14] |
Oda T, Wals P, Osterburg HH, et al. (1995) Clusterin (apoJ) alters the aggregation of amyloid β peptide (Aβ(1-42)) and forms slowly sedimenting Aβ complexes that cuase oxidative stress. Exp Neurol 136: 22–31. doi: 10.1006/exnr.1995.1080
|
| [15] |
Esparza TJ, Zhao H, Cirrito JR, et al. (2013) Amyloid-β oligomerization in Alzheimer dementia versus high-pathology controls. Ann Neurol 73: 104–119. doi: 10.1002/ana.23748
|
| [16] |
Gong YS, Chang L, Viola KL, et al. (2003) Alzheimer's disease-affected brain: Presence of oligomeric A β ligands (ADDLs) suggests a molecular basis for reversible memory loss. P Natl Acad Sci USA 100: 10417–10422. doi: 10.1073/pnas.1834302100
|
| [17] |
Kayed R, Head E, Thompson JL, et al. (2003) Common structure of soluble amyloid oligomers implies common mechanism of pathogenesis. Science 300: 486–489. doi: 10.1126/science.1079469
|
| [18] |
Noguchi A, Matsumura S, Dezawa M, et al. (2009) Isolation and characterization of patient-derived, toxic, high mass amyloid β-protein (Aβ) assembly from Alzheimer disease brains. J Biol Chem 284: 32895–32905. doi: 10.1074/jbc.M109.000208
|
| [19] |
Pham E, Crews L, Ubhi K, et al. (2010) Progressive accumulation of amyloid-β oligomers in Alzheimer's disease and in amyloid precursor protein transgenic mice is accompanied by selective alterations in synaptic scaffold proteins. FEBS J 277: 3051–3067. doi: 10.1111/j.1742-4658.2010.07719.x
|
| [20] | Gyure KA, Durham R, Stewart WF, et al. (2001) Intraneuronal Aβ-amyloid precedes development of amyloid plaques in Down syndrome. Arch Pathol Lab Med 125: 489–492. |
| [21] |
Lacor PN, Buniel MC, Chang L, et al. (2004) Synaptic targeting by Alzheimer's-related amyloid β oligomers. J Neurosci 24: 10191–10200. doi: 10.1523/JNEUROSCI.3432-04.2004
|
| [22] |
Lesne SE, Sherman MA, Grant M, et al. (2013) Brain amyloid-β oligomers in ageing and Alzheimer's disease. Brain 136: 1383–1398. doi: 10.1093/brain/awt062
|
| [23] |
Georganopoulou DG, Chang L, Nam JM, et al. (2005) Nanoparticle-based detection in cerebral spinal fluid of a soluble pathogenic biomarker for Alzheimer's disease. P Natl Acad Sci USA 102: 2273–2276. doi: 10.1073/pnas.0409336102
|
| [24] |
Bruggink KA, Jongbloed W, Biemans EALM, et al. (2013) Amyloid-β oligomer detection by ELISA in cerebrospinal fluid and brain tissue. Anal Biochem 433: 112–120. doi: 10.1016/j.ab.2012.09.014
|
| [25] |
Englund H, Gunnarsson MD, Brundin RM, et al. (2009) Oligomerization partially explains the lowering of Aβ42 in Alzheimer's disease cerebrospinal fluid. Neurodegener Dis 6: 139–147. doi: 10.1159/000225376
|
| [26] |
Fukumoto H, Tokuda T, Kasai T, et al. (2010) High-molecular-weight β-amyloid oligomers are elevated in cerebrospinal fluid of Alzheimer patients. FASEB J 24: 2716–2726. doi: 10.1096/fj.09-150359
|
| [27] |
Gao CM, Yam AY, Wang X, et al. (2010) Aβ40 Oligomers identified as a potential biomarker for the diagnosis of Alzheimer's disease. PLoS One 5: e15725. doi: 10.1371/journal.pone.0015725
|
| [28] |
Herskovits AZ, Locascio JJ, Peskind ER, et al. (2013) A luminex assay detects amyloid-β oligomers in Alzheimer's disease cerebrospinal fluid. PLoS One 8: e67898. doi: 10.1371/journal.pone.0067898
|
| [29] |
Holtta M, Hansson O, Andreasson U, et al. (2013) Evaluating amyloid-β oligomers in cerebrospinal fluid as a biomarker for Alzheimer's disease. PLoS One 8: e66381. doi: 10.1371/journal.pone.0066381
|
| [30] |
Jongbloed W, Bruggink KA, Kester MI, et al. (2015) Amyloid-β oligomers relate to cognitive decline in Alzheimer's disease. J Alzheimers Dis 45: 35–43. doi: 10.3233/JAD-142136
|
| [31] |
Santos AN, Ewers M, Minthon L, et al. (2012) Amyloid-β oligomers in cerebrospinal fluid are associated with cognitive decline in patients with Alzheimer's disease. J Alzheimers Dis 29: 171–176. doi: 10.3233/JAD-2012-111361
|
| [32] |
Lesne S, Koh MT, Kotilinek L, et al. (2006) A specific amyloid-β protein assembly in the brain impairs memory. Nature 440: 352–357. doi: 10.1038/nature04533
|
| [33] |
Ono K, Condron MM, Teplow DB (2009) Structure-neurotoxicity relationships of amyloid β-protein oligomers. P Natl Acad Sci USA 106: 14745–14750. doi: 10.1073/pnas.0905127106
|
| [34] |
Quist A, Doudevski L, Lin H, et al. (2005) Amyloid ion channels: A common structural link for protein-misfolding disease. P Natl Acad Sci USA 102: 10427–10432. doi: 10.1073/pnas.0502066102
|
| [35] |
Shankar GM, Bloodgood BL, Townsend M, et al. (2007) Natural oligomers of the Alzheimer amyloid-β protein induce reversible synapse loss by modulating an NMDA-type glutamate receptor-dependent signaling pathway. J Neurosci 27: 2866–2875. doi: 10.1523/JNEUROSCI.4970-06.2007
|
| [36] |
Shankar GM, Li S, Mehta TH, et al. (2008) Amyloid-β protein dimers isolated directly from Alzheimer's brains impair synaptic plasticity and memory. Nat Med 14: 837–842. doi: 10.1038/nm1782
|
| [37] |
Townsend M, Shankar GM, Mehta T, et al. (2006) Effects of secreted oligomers of amyloid β-protein on hippocampal synaptic plasticity: A potent role for trimers. J Physiol 572: 477–492. doi: 10.1113/jphysiol.2005.103754
|
| [38] |
Lambert MP, Barlow AK, Chromy BA, et al. (1998) Diffusible, nonfibrillar ligands derived from Aβ1-42 are potent central nervous system neurotoxins. P Natl Acad Sci USA 95: 6448–6453. doi: 10.1073/pnas.95.11.6448
|
| [39] |
Walsh DM, Klyubin I, Fadeeva JV, et al. (2002) Naturally secreted oligomers of amyloid β protein potently inhibit hippocampal long-term potentiation in vivo. Nature 416: 535–539. doi: 10.1038/416535a
|
| [40] |
Zhao J, Li A, Rajsombath M, et al. (2018) Soluble Aβ Oligomers Impair Dipolar Heterodendritic Plasticity by Activation of mGluR in the Hippocampal CA1 Region. iScience 6: 138–150. doi: 10.1016/j.isci.2018.07.018
|
| [41] |
De Felice FG, Wu D, Lambert MP, et al. (2008) Alzheimer's disease-type neuronal tau hyperphosphorylation induced by Aβ oligomers. Neurobiol Aging 29: 1334–1347. doi: 10.1016/j.neurobiolaging.2007.02.029
|
| [42] |
Ma QL, Yang F, Rosario ER, et al. (2009) β-amyloid oligomers induce phosphorylation of tau and inactivation of insulin receptor substrate via c-Jun N-terminal kinase signaling: Suppression by omega-3 fatty acids and curcumin. J Neurosci 29: 9078–9089. doi: 10.1523/JNEUROSCI.1071-09.2009
|
| [43] |
Resende R, Ferreiro E, Pereira C, et al. (2008) ER stress is involved in Aβ-induced GSK-3β activation and tau phosphorylation. J Neurosci Res 86: 2091–2099. doi: 10.1002/jnr.21648
|
| [44] |
Tomiyama T, Matsuyama S, Iso H, et al. (2010) A mouse model of amyloid-β oligomers: their contribution to synaptic alteration, abnormal tau phosphorylation, glial activation, and neuronal loss in vivo. J Neurosci 30: 4845–4856. doi: 10.1523/JNEUROSCI.5825-09.2010
|
| [45] |
Zempel H, Thies E, Mandelkow E, et al. (2010) A Oligomers Cause Localized Ca2+ Elevation, Missorting of Endogenous Tau into Dendrites, Tau Phosphorylation, and Destruction of Microtubules and Spines. J Neurosci 30: 11938–11950. doi: 10.1523/JNEUROSCI.2357-10.2010
|
| [46] |
Heinitz K, Beck M, Schliebs R, et al. (2006) Toxicity mediated by soluble oligomers of β-amyloid(1‑42) on cholinergic SN56.B5.G4 cells. J Neurochem 98: 1930–1945. doi: 10.1111/j.1471-4159.2006.04015.x
|
| [47] |
Nunes-Tavares N, Santos LE, Stutz B, et al. (2012) Inhibition of choline acetyltransferase as a mechanism for cholinergic dysfunction induced by amyloid-β peptide oligomers. J Biol Chem 287: 19377–19385. doi: 10.1074/jbc.M111.321448
|
| [48] |
De Felice FG, Velasco PT, Lambert MP, et al. (2007) Aβ oligomers induce neuronal oxidative stress through an N-methyl-D-aspartate receptor-dependent mechanism that is blocked by the Alzheimer drug memantine. J Biol Chem 282: 11590–11601. doi: 10.1074/jbc.M607483200
|
| [49] | Longo VD, Viola KL, Klein WL, et al. (2000) Reversible inactivation of superoxide-sensitive aconitase in Aβ1-42-treated neuronal cell lines. J Neurochem 75: 1977–1985. |
| [50] |
Sponne I, Fifre A, Drouet B, et al. (2003) Apoptotic neuronal cell death induced by the non-fibrillar amyloid-β peptide proceeds through an early reactive oxygen species-dependent cytoskeleton perturbation. J Biol Chem 278: 3437–3445. doi: 10.1074/jbc.M206745200
|
| [51] |
Tabner BJ, El-Agnaf OMA, Turnbull S, et al. (2005) Hydrogen peroxide is generated during the very early stages of aggregation of the amyloid peptides implicated in Alzheimer disease and familial British dementia. J Biol Chem 280: 35789–35792. doi: 10.1074/jbc.C500238200
|
| [52] |
Alberdi E, Wyssenbach A, Alberdi M, et al. (2013) Ca2+-dependent endoplasmic reticulum stress correlates with astrogliosis in oligomeric amyloid β-treated astrocytes and in a model of Alzheimer's disease. Aging Cell 12: 292–302. doi: 10.1111/acel.12054
|
| [53] |
Nishitsuji K, Tomiyama T, Ishibashi K, et al. (2009) The E693Δ mutation in amyloid precursor protein increases intracellular accumulation of amyloid β oligomers and causes endoplasmic reticulum stress-induced apoptosis in cultured cells. Am J Pathol 174: 957–969. doi: 10.2353/ajpath.2009.080480
|
| [54] |
Lacor PN, Buniel MC, Furlow PW, et al. (2007) Aβ oligomer-induced aberrations in synapse composition, shape, and density provide a molecular basis for loss of connectivity in Alzheimer's disease. J Neurosci 27: 796–807. doi: 10.1523/JNEUROSCI.3501-06.2007
|
| [55] |
Roselli F (2005) Soluble β-Amyloid1-40 induces NMDA-dependent degradation of postsynaptic density-95 at glutamatergic synapses. J Neurosci 25: 11061–11070. doi: 10.1523/JNEUROSCI.3034-05.2005
|
| [56] |
Snyder EM, Nong Y, Almeida CG, et al. (2005) Regulation of NMDA receptor trafficking by amyloid-β. Nat Neurosci 8: 1051–1058. doi: 10.1038/nn1503
|
| [57] | Zhao WQ, De Felice FG, Fernandez S, et al. (2007) Amyloid β oligomers induce impairment of neuronal insulin receptors. FASEB J 22: 246–260. |
| [58] |
De Felice FG, Vieira MN, Bomfim TR, et al. (2009) Protection of synapses against Alzheimer's-linked toxins: insulin signaling prevents the pathogenic binding of Aβ oligomers. Proc Natl Acad Sci USA 106: 1971–1976. doi: 10.1073/pnas.0809158106
|
| [59] |
Koffie RM, Meyer-Luehmann M, Hashimoto T, et al. (2009) Oligomeric amyloid-β associates with postsynaptic densities and correlates with excitatory synapse loss near senile plaques. P Natl Acad Sci USA 106: 4012–4017. doi: 10.1073/pnas.0811698106
|
| [60] |
Decker H, Lo KY, Unger SM, et al. (2010) Amyloid-Peptide oligomers disrupt axonal transport through an NMDA receptor-dependent mechanism that is mediated by glycogen synthase kinase 3 in primary cultured hippocampal neurons. J Neurosci 30: 9166–9171. doi: 10.1523/JNEUROSCI.1074-10.2010
|
| [61] |
Pigino G, Morfini G, Atagi Y, et al. (2009) Disruption of fast axonal transport is a pathogenic mechanism for intraneuronal amyloid β. P Natl Acad Sci USA 106: 5907–5912. doi: 10.1073/pnas.0901229106
|
| [62] |
Poon WW, Blurton-Jones M, Tu CH, et al. (2011) β-Amyloid impairs axonal BDNF retrograde trafficking. Neurobiol Aging 32: 821–833. doi: 10.1016/j.neurobiolaging.2009.05.012
|
| [63] |
Hu J, Akama KT, Krafft GA, et al. (1998) Amyloid-β peptide activates cultured astrocytes: Morphological alterations, cytokine induction and nitric oxide release. Brain Res 785: 195–206. doi: 10.1016/S0006-8993(97)01318-8
|
| [64] |
Jimenez S, Baglietto-Vargas D, Caballero C, et al. (2008) Inflammatory response in the hippocampus of PS1M146L/APP751SL mouse model of Alzheimer's disease: Age-dependent switch in the microglial phenotype from alternative to classic. J Neurosci 28: 11650–11661. doi: 10.1523/JNEUROSCI.3024-08.2008
|
| [65] |
Bhaskar K, Miller M, Chludzinski A, et al. (2009) The PI3K-Akt-mTOR pathway regulates a oligomer induced neuronal cell cycle events. Mol Neurodegener 4: 1–18. doi: 10.1186/1750-1326-4-1
|
| [66] |
Varvel NH, Bhaskar K, Patil AR, et al. (2008) Aβ oligomers induce neuronal cell cycle events in Alzheimer's disease. J Neurosci 28: 10786–10793. doi: 10.1523/JNEUROSCI.2441-08.2008
|
| [67] |
Kim HJ, Chae SC, Lee DK, et al. (2003) Selective neuronal degeneration induced by soluble oligomeric amyloid β protein. FASEB J 17: 118–120. doi: 10.1096/fj.01-0987fje
|
| [68] |
Roher AE, Lowenson JD, Clarke S, et al. (1993) β-Amyloid-(1-42) is a major component of cerebrovascular amyloid deposits: implications for the pathology of Alzheimer disease. P Natl Acad Sci USA 90: 10836. doi: 10.1073/pnas.90.22.10836
|
| [69] |
Näslund J, Schierhorn A, Hellman U, et al. (1994) Relative abundance of Alzheimer Aβ amyloid peptide variants in Alzheimer disease and normal aging. P Natl Acad Sci USA 91: 8378. doi: 10.1073/pnas.91.18.8378
|
| [70] |
Vandersteen A, Hubin E, Sarroukh R, et al. (2012) A comparative analysis of the aggregation behavior of amyloid-β peptide variants. FEBS Lett 586: 4088–4093. doi: 10.1016/j.febslet.2012.10.022
|
| [71] |
Barrow CJ, Yasuda A, Kenny PTM, et al. (1992) Solution conformations and aggregational properties of synthetic amyloid β-peptides of Alzheimer's disease: Analysis of circular dichroism spectra. J Mol Biol 225: 1075–1093. doi: 10.1016/0022-2836(92)90106-T
|
| [72] |
Barrow CJ, Zagorski MG (1991) Solution structures of β peptide and its constituent fragments: Relation to amyloid deposition. Science 253: 179–182. doi: 10.1126/science.1853202
|
| [73] |
Inouye H, Fraser PE, Kirschner DA (1993) Structure of β-crystallite assemblies formed by Alzheimer β-amyloid protein analogs-analysis by x-ray diffraction. Biophys J 64: 502–519. doi: 10.1016/S0006-3495(93)81393-6
|
| [74] |
Torok M, Milton S, Kayed R, et al. (2002) Structural and dynamic features of Alzheimer's Aβ peptide in amyloid fibrils studied by site-directed spin labeling. J Biol Chem 277: 40810–40815. doi: 10.1074/jbc.M205659200
|
| [75] |
Antzutkin ON, Balbach JJ, Leapman RD, et al. (2000) Multiple quantum solid-state NMR indicates a parallel, not antiparallel, organization of β-sheets in Alzheimer's β-amyloid fibrils. P Natl Acad Sci USA 97: 13045–13050. doi: 10.1073/pnas.230315097
|
| [76] |
Petkova AT, Leapman RD, Guo ZH, et al. (2005) Self-propagating, molecular-level polymorphism in Alzheimer's β-amyloid fibrils. Science 307: 262–265. doi: 10.1126/science.1105850
|
| [77] |
Tycko R (2006) Solid-state NMR as a probe of amyloid structure. Protein Peptide Lett 13: 229–234. doi: 10.2174/092986606775338470
|
| [78] |
Pastor MT, Kuemmerer N, Schubert V, et al. (2008) Amyloid toxicity is independent of polypeptide sequence, length and chirality. J Mol Biol 375: 695–707. doi: 10.1016/j.jmb.2007.08.012
|
| [79] |
Chiti F, Dobson CM (2006) Protein misfolding, functional amyloid, and human disease. Annu Rev Biochem 75: 333–366. doi: 10.1146/annurev.biochem.75.101304.123901
|
| [80] |
Xue WF, Homans SW, Radford SE (2008) Systematic analysis of nucleation-dependent polymerization reveals new insights into the mechanism of amyloid self-assembly. Proc Natl Acad Sci USA 105: 8926–8931. doi: 10.1073/pnas.0711664105
|
| [81] |
Murphy R (2007) Kinetics of amyloid formation and membrane interaction with amyloidogenic proteins. BBA-Biomembranes 1768: 1923–1934. doi: 10.1016/j.bbamem.2006.12.014
|
| [82] |
Ghosh P, Vaidya A, Kumar A, et al. (2016) Determination of critical nucleation number for a single nucleation amyloid-β aggregation model. Math Biosci 273: 70–79. doi: 10.1016/j.mbs.2015.12.004
|
| [83] |
Garai K, Sahoo B, Sengupta P, et al. (2008) Quasihomogeneous nucleation of amyloid β yields numerical bounds for the critical radius, the surface tension, and the free energy barrier for nucleus formation. J Chem Phys 128: 045102. doi: 10.1063/1.2822322
|
| [84] |
Novo M, Freire S, Al-Soufi W (2018) Critical aggregation concentration for the formation of early Amyloid-β(1-42) oligomers. Sci Rep 8: 1783. doi: 10.1038/s41598-018-19961-3
|
| [85] |
Xue C, Lin TY, Chang D, et al. (2017) Thioflavin T as an amyloid dye: Fibril quantification, optimal concentration and effect on aggregation. Roy Soc Open Sci 4: 160696. doi: 10.1098/rsos.160696
|
| [86] |
Kodali R, Wetzel R (2007) Polymorphism in the intermediates and products of amyloid assembly. Curr Opin Struc Biol 17: 48–57. doi: 10.1016/j.sbi.2007.01.007
|
| [87] |
Kodali R, Williams AD, Chemuru S, et al. (2010) Aβ(1-40) forms five distinct amyloid structures whose β-sheet contents and fibril stabilities are correlated. J Mol Biol 401: 503–517. doi: 10.1016/j.jmb.2010.06.023
|
| [88] |
Meinhardt J, Sachse C, Hortschansky P, et al. (2009) Aβ(1-40) fibril polymorphism implies diverse interaction patterns in amyloid fibrils. J Mol Biol 386: 869–877. doi: 10.1016/j.jmb.2008.11.005
|
| [89] |
Tycko R (2015) Amyloid polymorphism: Structural basis and neurobiological relevance. Neuron 86: 632–645. doi: 10.1016/j.neuron.2015.03.017
|
| [90] |
Colletier JP, Laganowsky A, Landau M, et al. (2011) Molecular basis for amyloid-β polymorphism. P Natl Acad Sci USA 108: 16938–16943. doi: 10.1073/pnas.1112600108
|
| [91] |
Crowther RA, Goedert M (2000) Abnormal Tau-containing filaments in Neurodegenerative Disease. J Struct Biol 130: 271–279. doi: 10.1006/jsbi.2000.4270
|
| [92] |
Lu JX, Qiang W, Yau WM, et al. (2013) Molecular structure of β-amyloid fibrils in Alzheimer's disease brain tissue. Cell 154: 1257–1268. doi: 10.1016/j.cell.2013.08.035
|
| [93] |
Qiang W, Yau WM, Lu JX, et al. (2017) Structural variation in amyloid-β fibrils from Alzheimer's disease clinical subtypes. Nature 541: 217–221. doi: 10.1038/nature20814
|
| [94] |
Paravastu AK, Qahwash I, Leapman RD, et al. (2009) Seeded growth of β-amyloid fibrils from Alzheimer's brain-derived fibrils produces a distinct fibril structure. P Natl Acad Sci USA 106: 7443–7448. doi: 10.1073/pnas.0812033106
|
| [95] |
Yates EA, Legleiter J (2014) Preparation protocols of Aβ(1-40) promote the formation of polymorphic aggregates and altered interactions with lipid bilayers. Biochemistry 53: 7038–7050. doi: 10.1021/bi500792f
|
| [96] |
Teplow DB (2013) On the subject of rigor in the study of amyloid β-protein assembly. Alzheimers Res Ther 5: 39. doi: 10.1186/alzrt203
|
| [97] |
Lee MC, Yu WC, Shih YH, et al. (2018) Zinc ion rapidly induces toxic, off-pathway amyloid-β oligomers distinct from amyloid-β derived diffusible ligands in Alzheimer's disease. Sci Rep 8: 1–16. doi: 10.1038/s41598-017-17765-5
|
| [98] |
Ryan DA, Narrow WC, Federoff HJ, et al. (2010) An improved method for generating consistent soluble amyloid-β oligomer preparations for in vitro neurotoxicity studies. J Neurosci Meth 190: 171–179. doi: 10.1016/j.jneumeth.2010.05.001
|
| [99] |
Barghorn S, Nimmrich V, Striebinger A, et al. (2005) Globular amyloid β-peptide1-42 oligomer-A homogenous and stable neuropathological protein in Alzheimer's disease. J Neurochem 95: 834–847. doi: 10.1111/j.1471-4159.2005.03407.x
|
| [100] |
Thibaudeau TA, Anderson RT, Smith DM (2018) A common mechanism of proteasome impairment by neurodegenerative disease-associated oligomers. Nat Commun 9: 1097. doi: 10.1038/s41467-018-03509-0
|
| [101] |
Stine WB, Dahlgren KN, Krafft GA, et al. (2003) In vitro characterization of conditions for amyloid-b peptide oligomerization and fibrillogenesis. J Biol Chem 278: 11612–11622. doi: 10.1074/jbc.M210207200
|
| [102] | Stine WB, Jungbauer L, Yu C, et al. (2011) Preparing synthetic Aβ in different aggregation states, In: Roberson ED (editor.), Alzheimer's Disease and Frontotemporal Dementia. Methods in Molecular Biology (Methods and Protocols), Totowa, NJ: Humana Press, 13–32. |
| [103] | Benninger RJ, David T (1983) An improved method of preparing the amyloid β-protein for fibrillogenesis and neurotoxicity experiments. Brain Res Rev 287: 173–196. |
| [104] |
Ryan TM, Caine J, Mertens HDT, et al. (2013) Ammonium hydroxide treatment of Aβ produces an aggregate free solution suitable for biophysical and cell culture characterization. PeerJ 1: e73. doi: 10.7717/peerj.73
|
| [105] |
Bitan G, Lomakin A, Teplow DB (2001) Amyloid β-protein oligomerization: Prenucleation interactions revealed by photo-induced cross-linking of unmodified proteins. J Biol Chem 276: 35176–35184. doi: 10.1074/jbc.M102223200
|
| [106] | Lesne SE (2013) Breaking the code of amyloid-β oligomers. Int J Cell Biol 2013: 950783. |
| [107] |
Sengupta U, Nilson AN, Kayed R (2016) The role of amyloid-β oligomers in toxicity, propagation, and immunotherapy. EBioMedicine 6: 42–49. doi: 10.1016/j.ebiom.2016.03.035
|
| [108] |
Benilova I, Karran E, De Strooper B (2012) The toxic Aβ oligomer and Alzheimer's disease: An emperor in need of clothes. Nat Neurosci 15: 349–357. doi: 10.1038/nn.3028
|
| [109] | Ferreira ST, Lourenco MV, Oliveira MM, et al. (2015) Soluble amyloid-β oligomers as synaptotoxins leading to cognitive impairment in Alzheimer's disease. Front Cell Neurosci 9: 191. |
| [110] |
Cline EN, Bicca MA, Viola KL, et al. (2018) The amyloid-β oligomer hypothesis: Beginning of the third decade. J Alzheimers Dis 64: S567–S610. doi: 10.3233/JAD-179941
|
| [111] |
Brody DL, Jiang H, Wildburger N, et al. (2017) Non-canonical soluble amyloid-β aggregates and plaque buffering: Controversies and future directions for target discovery in Alzheimer's disease. Alzheimers Res Ther 9: 62. doi: 10.1186/s13195-017-0293-3
|
| [112] |
Sherman MA, LaCroix M, Amar F, et al. (2016) Soluble conformers of Aβ and Tau alter selective proteins governing axonal transport. J Neurosci 36: 9647–9658. doi: 10.1523/JNEUROSCI.1899-16.2016
|
| [113] |
O'Malley TT, Oktaviani NA, Zhang D, et al. (2014) Aβ dimers differ from monomers in structural propensity, aggregation paths and population of synaptotoxic assemblies. Biochem J 461: 413–426. doi: 10.1042/BJ20140219
|
| [114] |
Cheng IH, Scearce-Levie K, Legleiter J, et al. (2007) Accelerating amyloid-β fibrillization reduces oligomer levels and functional deficits in Alzheimer disease mouse models. J Biol Chem 282: 23818–23828. doi: 10.1074/jbc.M701078200
|
| [115] |
Amar F, Sherman MA, Rush T, et al. (2017) The amyloid-β oligomer Aβ*56 induces specific alterations in neuronal signaling that lead to tau phosphorylation and aggregation. Sci Signal 10: eaal2021. doi: 10.1126/scisignal.aal2021
|
| [116] |
Liu P, Reed MN, Kotilinek LA, et al. (2015) Quaternary structure defines a large class of amyloid-β oligomers neutralized by sequestration. Cell Rep 11: 1760–1771. doi: 10.1016/j.celrep.2015.05.021
|
| [117] |
Knight EM, Kim SH, Kottwitz JC, et al. (2016) Effective anti-Alzheimer Aβ therapy involves depletion of specific Aβ oligomer subtypes. Neurol Neuroimmunol Neuroinflamm 3: e237. doi: 10.1212/NXI.0000000000000237
|
| [118] |
Velasco PT, Heffern MC, Sebollela A, et al. (2012) Synapse-binding subpopulations of Aβ oligomers sensitive to peptide assembly blockers and scFv antibodies. ACS Chem Neurosci 3: 972–981. doi: 10.1021/cn300122k
|
| [119] |
Ryan TM, Roberts BR, McColl G, et al. (2015) Stabilization of nontoxic Aβ-oligomers: Insights into the mechanism of action of hydroxyquinolines in Alzheimer's disease. J Neurosci 35: 2871–2884. doi: 10.1523/JNEUROSCI.2912-14.2015
|
| [120] |
Ferreira IL, Ferreiro E, Schmidt J, et al. (2015) Aβ and NMDAR activation cause mitochondrial dysfunction involving ER calcium release. Neurobiol Aging 36: 680–692. doi: 10.1016/j.neurobiolaging.2014.09.006
|
| [121] |
Barz B, Liao Q, Strodel B (2018) Pathways of amyloid-β aggregation depend on oligomer shape. J Am Chem Soc 140: 319–327. doi: 10.1021/jacs.7b10343
|
| [122] |
Brito-Moreira J, Lourenco MV, Oliveira MM, et al. (2017) Interaction of amyloid-β (Aβ) oligomers with neurexin 2 and neuroligin 1 mediates synapse damage and memory loss in mice. J Biol Chem 292: 7327–7337. doi: 10.1074/jbc.M116.761189
|
| [123] |
Figueiredo CP, Clarke JR, Ledo JH, et al. (2013) Memantine rescues transient cognitive impairment caused by high-molecular-weight Aβ oligomers but not the persistent impairment induced by low-molecular-weight oligomers. J Neurosci 33: 9626–9317. doi: 10.1523/JNEUROSCI.0482-13.2013
|
| [124] |
Upadhaya AR, Lungrin I, Yamaguchi H, et al. (2012) High-molecular weight Aβ oligomers and protofibrils are the predominant Aβ species in the native soluble protein fraction of the AD brain. J Cell Mol Med 16: 287–295. doi: 10.1111/j.1582-4934.2011.01306.x
|
| [125] |
Mc Donald JM, O'Malley TT, Liu W, et al. (2015) The aqueous phase of Alzheimer's disease brain contains assemblies built from similar to 4 and similar to 7 kDa Aβ species. Alzheimers Dement 11: 1286–1305. doi: 10.1016/j.jalz.2015.01.005
|
| [126] |
Savioz A, Giannakopoulos P, Herrmann FR, et al. (2016) A study of Aβ oligomers in the temporal cortex and cerebellum of patients with neuropathologically confirmed Alzheimer's disease compared to aged controls. Neurodegener Dis 16: 398–406. doi: 10.1159/000446283
|
| [127] |
Breydo L, Kurouski D, Rasool S, et al. (2016) Structural differences between amyloid β oligomers. Biochem Bioph Res Co 477: 700–705. doi: 10.1016/j.bbrc.2016.06.122
|
| [128] |
Watanabe-Nakayama T, Ono K, Itami M, et al. (2016) High-speed atomic force microscopy reveals structural dynamics of amyloid β(1-42) aggregates. P Natl Acad Sci USA 113: 5835–5840. doi: 10.1073/pnas.1524807113
|
| [129] |
Matsumura S, Shinoda K, Yamada M, et al. (2011) Two distinct amyloid β-protein (Aβ) assembly pathways leading to oligomers and fibrils identified by combined fluorescence correlation spectroscopy, morphology, and toxicity analyses. J Biol Chem 286: 11555–11562. doi: 10.1074/jbc.M110.181313
|
| [130] |
Miller Y, Ma B, Nussinov R (2010) Polymorphism in Alzheimer Aβ amyloid organization reflects conformational selection in a rugged energy landscape. Chem Rev 110: 4820–4838. doi: 10.1021/cr900377t
|
| [131] |
Bernstein SL, Dupuis NF, Lazo ND, et al. (2009) Amyloid-β protein oligomerization and the importance of tetramers and dodecamers in the aetiology of Alzheimer's disease. Nat Chem 1: 326–331. doi: 10.1038/nchem.247
|
| [132] |
Economou NJ, Giammona MJ, Do TD, et al. (2016) Amyloid β-protein assembly and Alzheimer's disease: Dodecamers of Aβ42, but not of Aβ40, seed fibril formation. J Am Chem Soc 138: 1772–1775. doi: 10.1021/jacs.5b11913
|
| [133] |
Shamitko-Klingensmith N, Boyd JW, Legleiter J (2016) Microtubule modification influences cellular response to amyloid-β exposure. AIMS Biophysics 3: 261–285. doi: 10.3934/biophy.2016.2.261
|
| [134] |
Yates EA, Cucco EM, Legleiter J (2011) Point mutations in Aβ induce polymorphic aggregates at liquid/solid interfaces. ACS Chem Neurosci 2: 294–307. doi: 10.1021/cn200001k
|
| [135] |
Yates EA, Owens SL, Lynch MF, et al. (2013) Specific domains of Aβ facilitate aggregation on and association with lipid bilayers. J Mol Biol 425: 1915–1933. doi: 10.1016/j.jmb.2013.03.022
|
| [136] |
Glabe CG (2008) Structural classification of toxic amyloid oligomers. J Biol Chem 283: 29639–29643. doi: 10.1074/jbc.R800016200
|
| [137] |
Chromy BA, Nowak RJ, Lambert MP, et al. (2003) Self-assembly of Aβ(1-42) into globular neurotoxins. Biochemistry 42: 12749–12760. doi: 10.1021/bi030029q
|
| [138] |
Glabe CG (2006) Common mechanisms of amyloid oligomer pathogenesis in degenerative disease. Neurobiol Aging 27: 570–575. doi: 10.1016/j.neurobiolaging.2005.04.017
|
| [139] |
Lee EB, Leng LZ, Zhang B, et al. (2006) Targeting amyloid-β peptide (Aβ) oligomers by passive immunization with a conformation-selective monoclonal antibody improves learning and memory in Aβ precursor protein (APP) transgenic mice. J Biol Chem 281: 4292–4299. doi: 10.1074/jbc.M511018200
|
| [140] |
Lambert MP, Velasco PT, Chang L, et al. (2007) Monoclonal antibodies that target pathological assemblies of Aβ. J Neurochem 100: 23–35. doi: 10.1111/j.1471-4159.2006.04157.x
|
| [141] |
Hayden EY, Conovaloff JL, Mason A, et al. (2017) Preparation of pure populations of covalently stabilized amyloid β-protein oligomers of specific sizes. Anal Biochem 518: 78–85. doi: 10.1016/j.ab.2016.10.026
|
| [142] |
Ono K, Li L, Takamura Y, et al. (2012) Phenolic compounds prevent amyloid β-protein oligomerization and synaptic dysfunction by site-specific binding. J Biol Chem 287: 14631–14643. doi: 10.1074/jbc.M111.325456
|
| [143] |
Bitan G, Kirkitadze MD, Lomakin A, et al. (2003) Amyloid β-protein (Aβ) assembly: Aβ40 and Aβ42 oligomerize through distinct pathways. P Natl Acad Sci USA 100: 330–335. doi: 10.1073/pnas.222681699
|
| [144] |
Al-Hilaly YK, Williams TL, Stewart-Parker M, et al. (2013) A central role for dityrosine crosslinking of Amyloid-β in Alzheimer's disease. Acta Neuropathol Commun 1: 83. doi: 10.1186/2051-5960-1-83
|
| [145] | Bush AI (2013) The metal theory of Alzheimer's disease. J Alzheimers Dis 33: S277–S281. |
| [146] |
Butterfield DA, Boyd-Kimball D (2018) Oxidative stress, amyloid-β peptide, and altered key molecular pathways in the pathogenesis and progression of Alzheimer's disease. J Alzheimers Dis 62: 1345–1367. doi: 10.3233/JAD-170543
|
| [147] |
Smith DP, Ciccotosto GD, Tew DJ, et al. (2007) Concentration dependent Cu2+ induced aggregation and dityrosine formation of the Alzheimer's disease amyloid-β peptide. Biochemistry 46: 2881–2891. doi: 10.1021/bi0620961
|
| [148] |
Ryan TM, Kirby N, Mertens HDT, et al. (2015) Small angle X-ray scattering analysis of Cu2+-induced oligomers of the Alzheimer's amyloid β peptide. Metallomics 7: 536–543. doi: 10.1039/C4MT00323C
|
| [149] |
Takano K, Endo S, Mukaiyama A, et al. (2006) Structure of amyloid β fragments in aqueous environments. FEBS J 273: 150–158. doi: 10.1111/j.1742-4658.2005.05051.x
|
| [150] |
Streltsov VA, Varghese JN, Masters CL, et al. (2011) Crystal structure of the amyloid-β p3 fragment provides a model for oligomer formation in Alzheimer's disease. J Neurosci 31: 1419–1426. doi: 10.1523/JNEUROSCI.4259-10.2011
|
| [151] |
Liu C, Sawaya MR, Cheng PN, et al. (2011) Characteristics of amyloid-related oligomers revealed by crystal structures of macrocyclic β-sheet mimics. J Am Chem Soc 133: 6736–6744. doi: 10.1021/ja200222n
|
| [152] |
Pham JD, Chim N, Goulding CW, et al. (2013) Structures of oligomers of a peptide from β-amyloid. J Am Chem Soc 135: 12460–12467. doi: 10.1021/ja4068854
|
| [153] |
Spencer RK, Li H, Nowick JS (2014) X-ray crystallographic structures of trimers and higher-order oligomeric sssemblies of a peptide derived from Aβ17–36. J Am Chem Soc 136: 5595–5598. doi: 10.1021/ja5017409
|
| [154] |
Bhatia R, Lin H, Lal R (2000) Fresh and nonfibrillar amyloid b protein(1-42) induces rapid cellular degeneration in aged human fibroblasts: Evidence for AbP-channel-mediated cellular toxicity. FASEB 14: 1233–1243. doi: 10.1096/fasebj.14.9.1233
|
| [155] |
Lin H, Bhatia R, Lal R (2001) Amyloid b protein forms ion channels: Implications for Alzheimer's disease pathophysiology. FASEB 15: 2433–2444. doi: 10.1096/fj.01-0377com
|
| [156] |
Lin H, Zhu YJ, Lal R (1999) Amyloid-b protein (1–40) forms calcium-permeable, Zn2+-sensitive channel in reconstituted lipid vesicles. Biochemistry 38: 11189–11196. doi: 10.1021/bi982997c
|
| [157] |
Rhee SK, Quist AP, Lal R (1998) Amyloid b protein-(1-42) forms calcium-permeable, Zn2+-sensitive channel. J Biol Chem 273: 13379–13382. doi: 10.1074/jbc.273.22.13379
|
| [158] |
Parbhu A, Lin H, Thimm J, et al. (2002) Imaging real-time aggregation of amyloid β protein (1-42) by atomic force microscopy. Peptides 23: 1265–1270. doi: 10.1016/S0196-9781(02)00061-X
|
| [159] | Legleiter J, (2011) Assessing Aβ aggregation state by atomic force microscopy, In: Roberson ED (editor.), Alzheimer's Disease and Frontotemporal Dementia: Methods and Protocols, 57–70. |
| [160] |
Kowalewski T, Holtzman DM (1999) In situ atomic force microscopy study of Alzheimer's β-amyloid peptide on different substrates: New insights into mechanism of β-sheet formation. Proc Natl Acad Sci USA 96: 3688–3693. doi: 10.1073/pnas.96.7.3688
|
| [161] |
Hane F, Drolle E, Gaikwad R, et al. (2011) Amyloid-β aggregation on model lipid membranes: An atomic force microscopy study. J Alzheimers Dis 26: 485–494. doi: 10.3233/JAD-2011-102112
|
| [162] |
Legleiter J, Fryer JD, Holtzman DM, et al. (2011) The modulating effect of mechanical changes in lipid bilayers caused by apoE-containing lipoproteins on Aβ induced membrane disruption. ACS Chem Neurosci 2: 588–599. doi: 10.1021/cn2000475
|
| [163] |
Yip CM, Elton EA, Darabie AA, et al. (2001) Cholesterol, a modulator of membrane-associated Ab-fibrillogenesis and neurotoxicity. J Mol Biol 311: 723–734. doi: 10.1006/jmbi.2001.4881
|
| [164] |
Yip CM, McLaurin J (2001) Amyloid-b assembly: A critical step in fibrillogensis and membrane disruption. Biophys J 80: 1359–1371. doi: 10.1016/S0006-3495(01)76109-7
|
| [165] |
Pifer PM, Yates EA, Legleiter J (2011) Point mutations in Aβ result in the formation of distinct polymorphic aggregates in the presence of lipid bilayers. PLoS One 6: e16248. doi: 10.1371/journal.pone.0016248
|
| [166] |
Burke KA, Yates EA, Legleiter J (2013) Amyloid-forming proteins alter the local mechanical properties of lipid membranes. Biochemistry 52: 808–817. doi: 10.1021/bi301070v
|
| [167] |
Hane F, Tran G, Attwood S, et al. (2013) Cu2+ affects amyloid-β (1-42) aggregation by increasing peptide-peptide binding forces. PLoS One 8: e59005. doi: 10.1371/journal.pone.0059005
|
| [168] |
Kim BH, Palermo NY, Lovas S, et al. (2011) Single-molecule atomic force microscopy force spectroscopy study of Aβ-40 interactions. Biochemistry 50: 5154–5162. doi: 10.1021/bi200147a
|
| [169] |
Banerjee S, Sun Z, Hayden EY, et al. (2017) Nanoscale dynamics of amyloid-β42 oligomers as revealed by high-speed atomic force microscopy. ACS Nano 11: 12202–12209. doi: 10.1021/acsnano.7b05434
|
| [170] |
Yang T, Li S, Xu H, et al. (2017) Large soluble oligomers of amyloid β-protein from Alzheimer brain are far less neuroactive than the smaller oligomers to which they dissociate. J Neurosci 37: 152–163. doi: 10.1523/JNEUROSCI.1698-16.2016
|
| [171] |
Ahmed M, Davis J, Aucoin D, et al. (2010) Structural conversion of neurotoxic amyloid-β(1-42) oligomers to fibrils. Nat Struct Mol Biol 17: 561–556. doi: 10.1038/nsmb.1799
|
| [172] |
Chimon S, Shaibat MA, Jones CR, et al. (2007) Evidence of fibril-like β-sheet structures in a neurotoxic amyloid intermediate of Alzheimer's β-amyloid. Nat Struct Mol Biol 14: 1157–1164. doi: 10.1038/nsmb1345
|
| [173] |
Stroud JC, Liu C, Teng PK, et al. (2012) Toxic fibrillar oligomers of amyloid-β have cross-β structure. P Natl Acad Sci USA 109: 7717–7722. doi: 10.1073/pnas.1203193109
|
| [174] |
Gu L, Liu C, Stroud JC, et al. (2014) Antiparallel triple-strand architecture for prefibrillar Aβ42 oligomers. J Biol Chem 289: 27300–27313. doi: 10.1074/jbc.M114.569004
|
| [175] |
Teoh CL, Su D, Sahu S, et al. (2015) Chemical fluorescent probe for detection of Aβ oligomers. J Am Chem Soc 137: 13503–13509. doi: 10.1021/jacs.5b06190
|
| [176] |
Jameson LP, Dzyuba SV (2013) Aza-BODIPY: Improved synthesis and interaction with soluble Aβ1-42 oligomers. Bioorg Med Chem Lett 23: 1732–1735. doi: 10.1016/j.bmcl.2013.01.065
|
| [177] |
Ono M, Watanabe H, Kimura H, et al. (2012) BODIPY-based molecular probe for imaging of cerebral β-amyloid plaques. ACS Chem Neurosci 3: 319–324. doi: 10.1021/cn3000058
|
| [178] |
Verwilst P, Kim HR, Seo J, et al. (2017) Rational design of in vivo Tau tangle-selective near-infrared fluorophores: expanding the BODIPY universe. J Am Chem Soc 139: 13393–13403. doi: 10.1021/jacs.7b05878
|
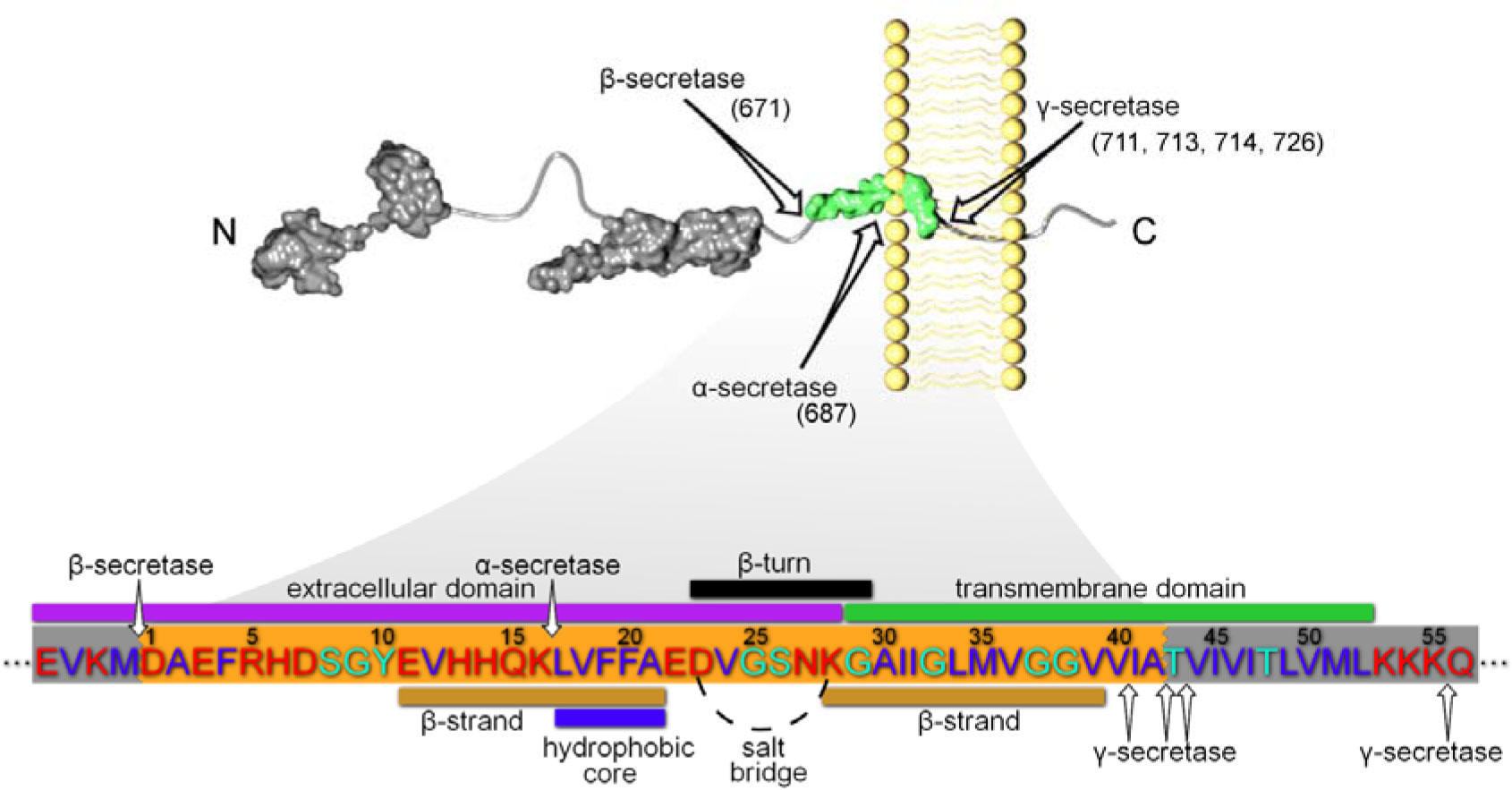
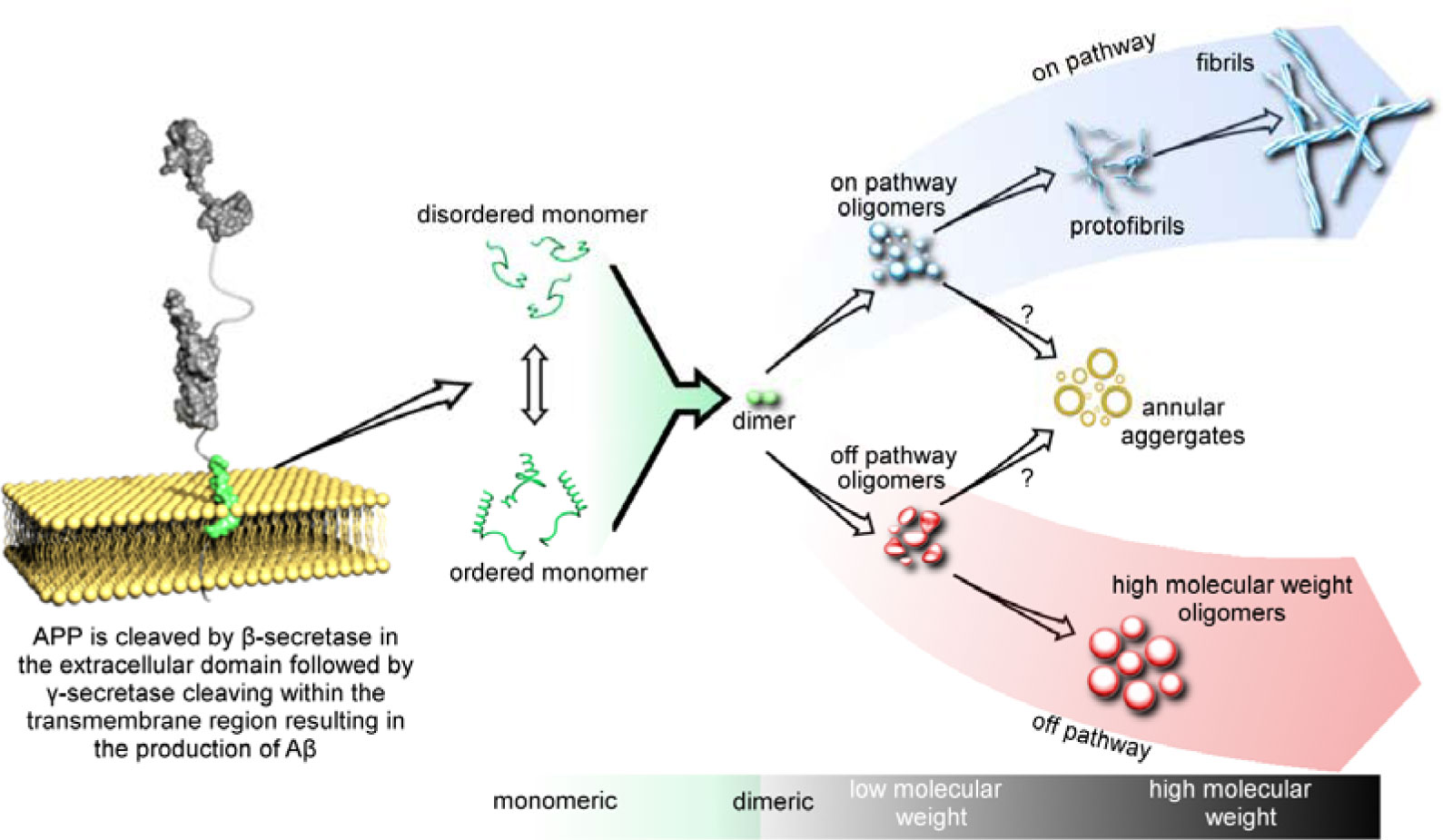
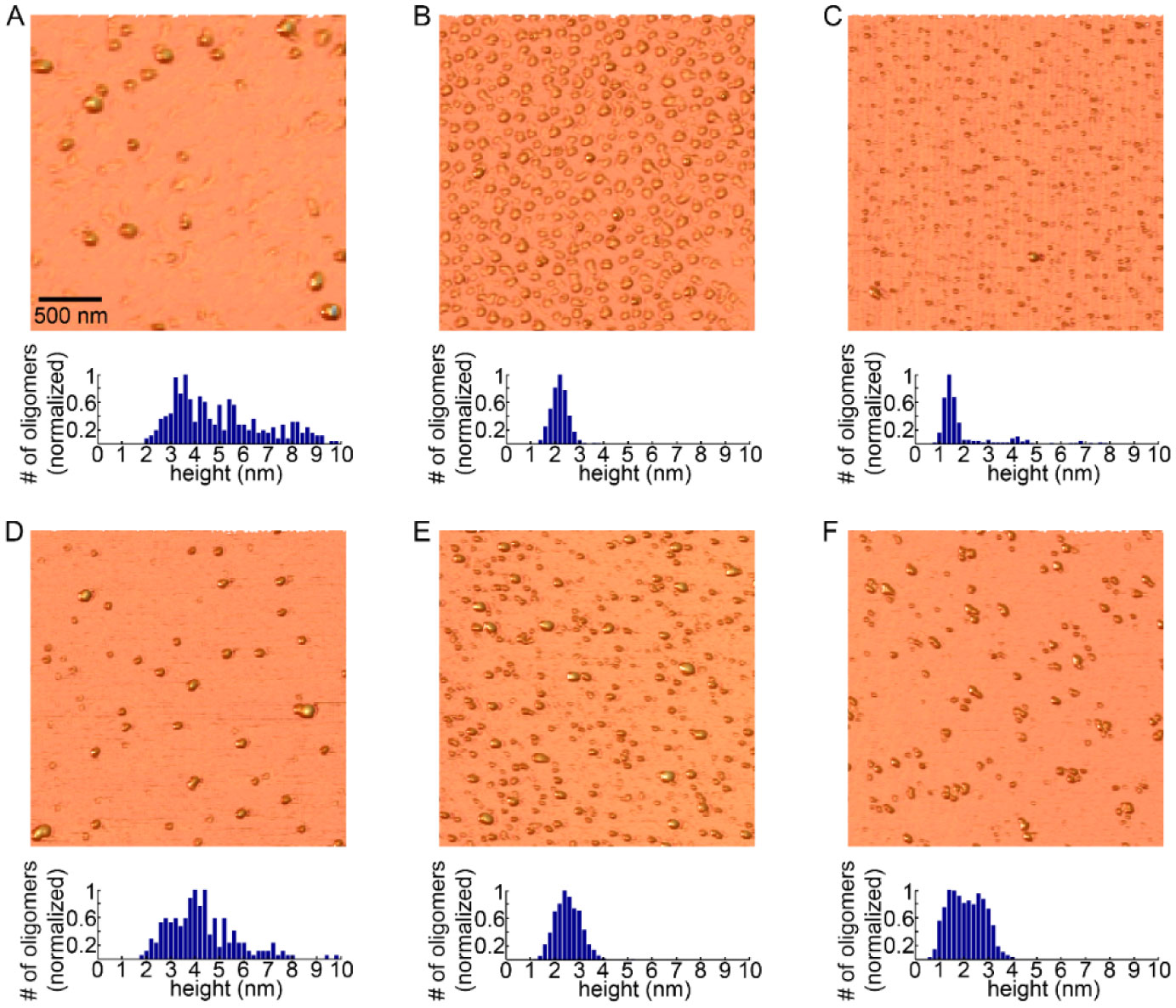
Figures(3) / Tables(4)

Albert W. Pilkington IV, Justin Legleiter. Challenges in understanding the structure/activity relationship of Aβ oligomers[J]. AIMS Biophysics, 2019, 6(1): 1-22. doi: 10.3934/biophy.2019.1.1







 DownLoad:
DownLoad: