Citation: Zachary Mekus, Jessica Cooley, Aaron George, Victoria Sabo, Morgan Strzegowski, Michelle Starz-Gaiano, Bradford E. Peercy. Effects of cell packing on chemoattractant distribution within a tissue[J]. AIMS Biophysics, 2018, 5(1): 1-21. doi: 10.3934/biophy.2018.1.1

| [1] |
Muller P, Schier AF (2011) Extracellular movement of signaling molecules. Dev Cell 21: 145–158. doi: 10.1016/j.devcel.2011.06.001

|
| [2] |
Briscoe J, Small S (2015) Morphogen rules: Design principles of gradient-mediated embryo patterning. Development 142: 3996–4009. doi: 10.1242/dev.129452
|
| [3] |
Kicheva A, Bollenbach T, Wartlick O, et al. (2012) Investigating the principles of morphogen gradient formation: From tissues to cells. Curr Opin Genet Dev 22: 527–532. doi: 10.1016/j.gde.2012.08.004
|
| [4] |
Janetopoulos C, Firtel RA (2008) Directional sensing during chemotaxis. FEBS Lett 582: 2075–2085. doi: 10.1016/j.febslet.2008.04.035
|
| [5] |
Roussos ET, Condeelis JS, Patsialou A (2011) Chemotaxis in cancer. Nat Rev Cancer 11: 573–587. doi: 10.1038/nrc3078
|
| [6] |
Cai D, Montell DJ (2014) Diverse and dynamic sources and sinks in gradient formation and directed migration. Curr Opin Cell Biol 30: 91–98. doi: 10.1016/j.ceb.2014.06.009
|
| [7] |
Haeger A, Wolf K, Zegers MM, et al. (2015) Collective cell migration: Guidance principles and hierarchies. Trends Cell Biol 25: 556–566. doi: 10.1016/j.tcb.2015.06.003
|
| [8] |
Danuser G, Allard J, Mogilner A (2013) Mathematical modeling of eukaryotic cell migration: Insights beyond experiments. Annu Rev Cell Dev Biol 29: 501–528. doi: 10.1146/annurev-cellbio-101512-122308
|
| [9] | Barua D, Parent SE, Winklbauer R (2017) Mechanics of Fluid-Filled Interstitial Gaps. II. Gap Characteristics in Xenopus Embryonic Ectoderm. Biophys J 113: 923–936. |
| [10] |
David R, Luu O, Damm EW, et al. (2014) Tissue cohesion and the mechanics of cell rearrangement. Development 141: 3672–3682. doi: 10.1242/dev.104315
|
| [11] | Nicholson C, Hrabetova S (2017) Brain extracellular space: The final frontier of neuroscience. Biophys J. |
| [12] |
Hudson AM, Cooley L (2014) Methods for studying oogenesis. Methods 68: 207–217. doi: 10.1016/j.ymeth.2014.01.005
|
| [13] | Duhart JC, Parsons TT, Raftery LA (2017) The repertoire of epithelial morphogenesis on display: Progressive elaboration of Drosophila egg structure. Mech Develop. |
| [14] |
Losick VP, Morris LX, Fox DT, et al. (2011) Drosophila stem cell niches: A decade of discovery suggests a unified view of stem cell regulation. Dev Cell 21: 159–171. doi: 10.1016/j.devcel.2011.06.018
|
| [15] |
Cheung LS, Schupbach T, Shvartsman SY (2011) Pattern formation by receptor tyrosine kinases: Analysis of the Gurken gradient in Drosophila oogenesis. Curr Opin Genet Dev 21: 719–725. doi: 10.1016/j.gde.2011.07.009
|
| [16] | King RC (1970) Ovarian development in Drosophila melanogaster. Q Rev Biol 44: 487–495. |
| [17] |
Montell DJ, Wan HY, Starz-Gaiano M (2012) Group choreography: Mechanisms orchestrating the collective movement of border cells. Nat Rev Mol Cell Biol 13: 631–645. doi: 10.1038/nrm3433
|
| [18] |
Saadin A, Starz-Gaiano M (2016) Circuitous genetic regulation governs a straightforward cell migration. Trends Genet 32: 660–673. doi: 10.1016/j.tig.2016.08.001
|
| [19] |
Duchek P, Somogyi K, Jekely G, et al. (2001) Guidance of cell migration by the Drosophila PDGF/VEGF receptor. Cell 107: 17–26. doi: 10.1016/S0092-8674(01)00502-5
|
| [20] |
McDonald JA, Pinheiro EM, Montell DJ (2003) PVF1, a PDGF/VEGF homolog, is sufficient to guide border cells and interacts genetically with Taiman. Development 130: 3469–3478. doi: 10.1242/dev.00574
|
| [21] |
Bianco A, Poukkula M, Cliffe A, et al. (2007) Two distinct modes of guidance signalling during collective migration of border cells. Nature 448: 362–365. doi: 10.1038/nature05965
|
| [22] |
Prasad M, Montell DJ (2007) Cellular and molecular mechanisms of border cell migration analyzed using time-lapse live-cell imaging. Dev Cell 12: 997–1005. doi: 10.1016/j.devcel.2007.03.021
|
| [23] |
Duchek P, Rorth P (2001) Guidance of cell migration by EGF receptor signaling during Drosophila oogenesis. Science 291: 131–133. doi: 10.1126/science.291.5501.131
|
| [24] |
McDonald JA, Pinheiro EM, Kadlec L, et al. (2006) Multiple EGFR ligands participate in guiding migrating border cells. Dev Biol 296: 94–103. doi: 10.1016/j.ydbio.2006.04.438
|
| [25] |
Wasserman JD, Freeman M (1998) An autoregulatory cascade of EGF receptor signaling patterns the Drosophila egg. Cell 95: 355–364. doi: 10.1016/S0092-8674(00)81767-5
|
| [26] |
Stonko DP, Manning L, Starz-Gaiano M, et al. (2015) A mathematical model of collective cell migration in a three-dimensional, heterogeneous environment. PLoS One 10: e0122799. doi: 10.1371/journal.pone.0122799
|
| [27] |
Cai D, Dai W, Prasad M, et al. (2016) Modeling and analysis of collective cell migration in an in vivo three-dimensional environment. Proc Natl Acad Sci USA 113: E2134–E2141. doi: 10.1073/pnas.1522656113
|
| [28] |
Yamao M, Naoki H, Ishii S (2011) Multi-cellular logistics of collective cell migration. PLoS One 6: e27950. doi: 10.1371/journal.pone.0027950
|
| [29] |
Manning LA, Weideman AM, Peercy BE, et al. (2015) Tissue landscape alters adjacent cell fates during Drosophila egg development. Nat Commun 6: 7356. doi: 10.1038/ncomms8356
|
| [30] |
Rorth P (2002) Initiating and guiding migration: Lessons from border cells. Trends Cell Biol 12: 325–331. doi: 10.1016/S0962-8924(02)02311-5
|
| [31] |
Grimm O, Coppey M, Wieschaus E (2010) Modelling the Bicoid gradient. Development 137: 2253–2264. doi: 10.1242/dev.032409
|
| [32] |
Muller P, Rogers KW, Yu SR, et al. (2013) Morphogen transport. Development 140: 1621–1638. doi: 10.1242/dev.083519
|
| [33] |
Kicheva A, Pantazis P, Bollenbach T, et al. (2007) Kinetics of morphogen gradient formation. Science 315: 521–525. doi: 10.1126/science.1135774
|
| [34] |
Zhou S, Lo WC, Suhalim JL, et al. (2012) Free extracellular diffusion creates the Dpp morphogen gradient of the Drosophila wing disc. Curr Biol 22: 668–675. doi: 10.1016/j.cub.2012.02.065
|
| [35] |
Wright VM, Vogt KL, Smythe E, et al. (2011) Differential activities of the Drosophila JAK/STAT pathway ligands Upd, Upd2 and Upd3. Cell Signal 23: 920–927. doi: 10.1016/j.cellsig.2011.01.020
|
| [36] |
Krall JA, Beyer EM, MacBeath G (2011) High- and low-affinity epidermal growth factor receptor-ligand interactions activate distinct signaling pathways. PLoS One 6: e15945. doi: 10.1371/journal.pone.0015945
|
| [37] |
Kong Q, Majeska RJ, Vazquez M (2011) Migration of connective tissue-derived cells is mediated by ultra-low concentration gradient fields of EGF. Exp Cell Res 317: 1491–1502. doi: 10.1016/j.yexcr.2011.04.003
|
| [38] |
Raja WK, Gligorijevic B, Wyckoff J, et al. (2010) A new chemotaxis device for cell migration studies. Integr Biol 2: 696–706. doi: 10.1039/c0ib00044b
|
| [39] |
Wang SJ, Saadi W, Lin F, et al. (2004) Differential effects of EGF gradient profiles on MDA-MB-231 breast cancer cell chemotaxis. Exp Cell Res 300: 180–189. doi: 10.1016/j.yexcr.2004.06.030
|
| [40] | Kammermeyer KL, Wadsworth SC (1987) Expression of Drosophila epidermal growth factor receptor homologue in mitotic cell populations. Development 100: 201–210. |
| [41] |
Iglesias PA, Devreotes PN (2012) Biased excitable networks: How cells direct motion in response to gradients. Curr Opin Cell Biol 24: 245–253. doi: 10.1016/j.ceb.2011.11.009
|
| [42] |
Prasad M, Jang AC, Starz-Gaiano M, et al. (2007) A protocol for culturing Drosophila melanogaster stage 9 egg chambers for live imaging. Nat Protoc 2: 2467–2473. doi: 10.1038/nprot.2007.363
|
| [43] |
Lusk JB, Lam VYW, Tolwinski NS (2017) Epidermal growth factor pathway signaling in Drosophila embryogenesis: Tools for understanding cancer. Cancers 9: 16. doi: 10.3390/cancers9020016
|
| [44] |
Stephens L, Milne L, Hawkins P (2008) Moving towards a better understanding of chemotaxis. Curr Biol 18: R485–R494. doi: 10.1016/j.cub.2008.04.048
|
| [45] |
Hughes-Alford SK, Lauffenburger DA (2012) Quantitative analysis of gradient sensing: Towards building predictive models of chemotaxis in cancer. Curr Opin Cell Biol 24: 284–291. doi: 10.1016/j.ceb.2012.01.001
|
| [46] |
Gregor T, Wieschaus EF, McGregor AP, et al. (2007) Stability and nuclear dynamics of the bicoid morphogen gradient. Cell 130: 141–152. doi: 10.1016/j.cell.2007.05.026
|
| [47] |
Venkiteswaran G, Lewellis SW, Wang J, et al. (2013) Generation and dynamics of an endogenous, self-generated signaling gradient across a migrating tissue. Cell 155: 674–687. doi: 10.1016/j.cell.2013.09.046
|
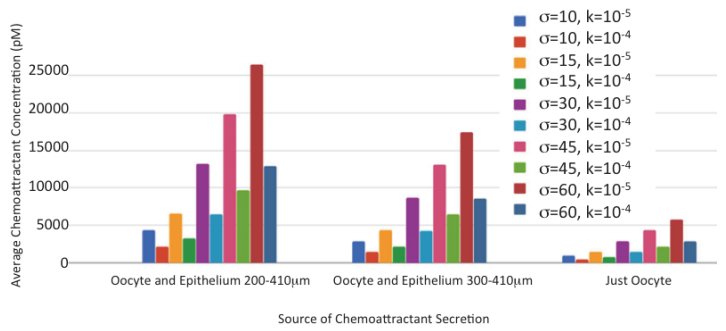
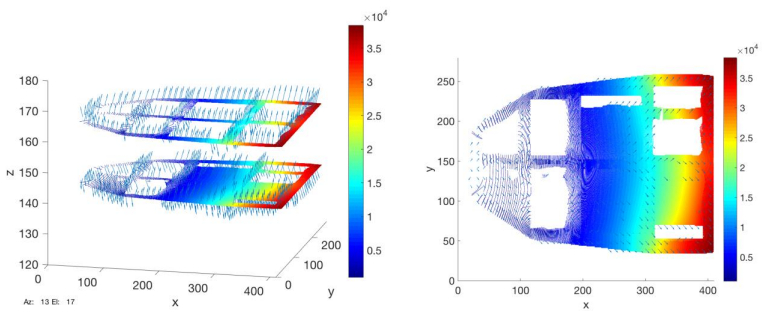
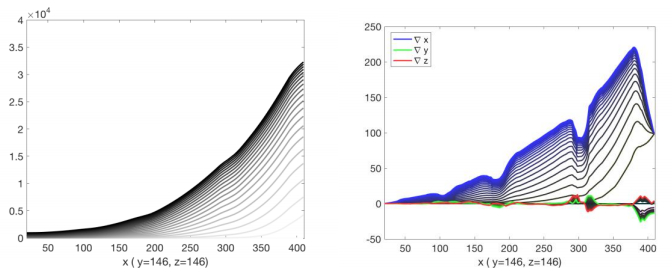
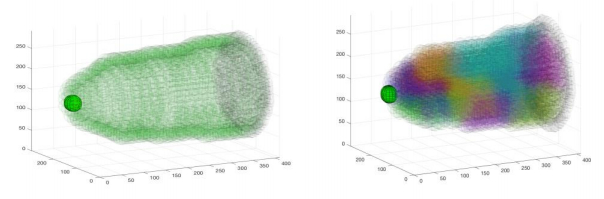
Figures(9) / Tables(7)

Zachary Mekus, Jessica Cooley, Aaron George, Victoria Sabo, Morgan Strzegowski, Michelle Starz-Gaiano, Bradford E. Peercy. Effects of cell packing on chemoattractant distribution within a tissue[J]. AIMS Biophysics, 2018, 5(1): 1-21. doi: 10.3934/biophy.2018.1.1







 DownLoad:
DownLoad: