Citation: Klemen Bohinc, Leo Lue. On the electrostatics of DNA in chromatin[J]. AIMS Biophysics, 2016, 3(1): 75-87. doi: 10.3934/biophy.2016.1.75

| [1] |
Korolev N, Vorontsova OV, Nordenskiold L (2007) Physicochemical analysis of electrostatic foundation for DNA-protein interactions in chromatin transformations. Prog Biophys Mol Biol 95: 23–49. doi: 10.1016/j.pbiomolbio.2006.11.003

|
| [2] | Bloomfield VA (1997) DNA condensation by multivalent cations. Biopolymers 44: 269. |
| [3] |
Teif VB, Bohinc K (2011) Physicochemical analysis of electrostatic foundation for DNA-protein interactions in chromatin transformations. Prog Biophys Mol Biol 105: 208–282. doi: 10.1016/j.pbiomolbio.2010.07.002
|
| [4] |
Radler JO, Koltover I, Salditt T, et al. (1997) Structure of DNA-cationic liposome complexes: DNA intercalation in multilamellar membranes in distinct interhelical packing regimes. Science 275: 810–814. doi: 10.1126/science.275.5301.810
|
| [5] |
Chow MH, Yan KTH, Bennett MJ, et al. (2010) Birefringence and DNA condensation of liquid crystalline chromosomes. Eukaryotic Cell 9:1577-1587. doi: 10.1128/EC.00026-10
|
| [6] | Gelbart WM, Bruinsma RF, Pincus PA, et al. (2000) DNA-inspired electrostatics Physicochemical analysis of electrostatic foundation for DNA-protein interactions in chromatin transformations. Physics Today 53:38–44. |
| [7] |
Mengistu DH, Bohinc K, May S, (2009) Binding of DNA to zwitterionic lipid layers mediated by divalent cations. J Phys Chem B 113: 12277–12282. doi: 10.1021/jp904986j
|
| [8] |
Raedler JO, Koltover I, Salditt T, et al. (1997) Structure of DNA-cationic liposome complexes: DNA intercalation in multilamellar membranes in distinct interhelical packing regimes. Science 275: 810–814. doi: 10.1126/science.275.5301.810
|
| [9] | Evans DF, Wennerström H (1994) The colloidal domain, where physics, chemistry, biology and technology meet. , 2 Eds., New York: VCH Publishers. |
| [10] |
Butler JC, Angelini T, Tang JX, et al. (2003) Ion multivalence and like-charge polyelectrolyte attraction. Phys Rev Lett 91: 028301. doi: 10.1103/PhysRevLett.91.028301
|
| [11] | Angelini TE, Liang H, Wriggers W, et al. (2003) Like-charge attraction between polyelectrolytes induced by counterion charge density waves. Proc Nat Acad Sci U S A 100: 8634–8637. |
| [12] | Bohinc K, Brezesinski G, May S (2012) Modeling the influence of adsorbed DNA on the lateral pressure and tilt transition of a zwitterionic lipid monolayer. Phys Chem Chem Phys 40: 10613–10621. |
| [13] | Gouy MG (1910) Sur la constitution dela charge electrique ala surface d‘un electrolyte. J Phys Radium (Paris) 9: 457–468. |
| [14] | Chapman DL (1913) A Contribution to the Theory of Electrocapillarity. Philos Mag 6: 455–481. |
| [15] |
Moreira AG, Netz RR (2001) Binding of similarly charged plates with counterions only Modeling the influence of adsorbed DNA on the lateral pressure and tilt transition of a zwitterionic lipid monolayer. Phys Rev Lett 87: 078301. doi: 10.1103/PhysRevLett.87.078301
|
| [16] | Shklovskii BI (1999) Screening of a macroion by multivalent ions: Correlation-induced inversion of charge. Phys Rev E 60: 5802–5811. |
| [17] |
Carnie S, McLaughlin S (1983) Large divalent-cations and electrostatic potentials adjacent to membranes - a theoretical calculation. Biophys J 44: 325–332. doi: 10.1016/S0006-3495(83)84306-9
|
| [18] | Kirkwood JG, Shumaker JB (1953) Forces Between Protein Molecules in Solution Arising from Fluctuations in Proton Charge and Configuration. Proc Nat Acad Sci U S A 38: 863–871. |
| [19] | Guldbrand L, Jönsson B, Wennerström H, et al. (1984) Electrical double layer forces. A Monte Carlo study. J Chem Phys 80: 2221–2228. |
| [20] | Reščič J, Linse P (2000) Charged colloidal solutions with short flexible counterions. J Phys Chem B 32: 7852–7857. |
| [21] | Svensson B, Jönsson B (1984) The interaction between charged aggregates in electrolyte solution - a Monte-Carlo simulation study. Chem Phys Lett 108: 580–584. |
| [22] |
Coalson RD, Duncan A (1992) Systematic ionic screening theory of macroions. J Chem Phys 97: 5653–5661. doi: 10.1063/1.463950
|
| [23] | Coalson RD, Walsh AM, Duncan A, et al. (1995) Statistical-mechanics of a Coulomb gas with finite-size particles - a lattice field theory. J Chem Phys 102: 4584–4594. |
| [24] | Tsonchev S, Coalson RD, Duncan A (1999) Statistical mechanics of charged polymers in electrolyte solutions: A lattice field theory approach. Phys Rev E 60: 4257–4267. |
| [25] |
Tsonchev S, Coalson RD, Duncan A (2007) Partitioning of a polymer chain between a confining cavity and a gel. Phys Rev E 76: 041804. doi: 10.1103/PhysRevE.76.041804
|
| [26] |
Navarre WW, Porwollik S, Wang Y, et al. (2006) Selective silencing of foreign DNA with low GC content by the H-NS protein in Salmonella. Science 313: 236–238. doi: 10.1126/science.1128794
|
| [27] | Bohinc K, Igliˇ c A, May S (2004) Interaction between macroions mediated by divalent rod-like ions. Europhys Lett 68: 494–500. |
| [28] |
May S, Igliˇ c S, Reščič S, et al. (2008) Bridging like-charged macroions through long divalent rodlike ions. J Phys Chem B 112: 1685–1692. doi: 10.1021/jp073355e
|
| [29] | Maset S, Bohinc K (2007) Orientations of dipoles restricted by two oppositely charged walls. J Phys A 40: 11815–11826. |
| [30] |
Maset S, Reščič J, May S, et al. (2009) Attraction between like-charged surfaces induced by orientational ordering of divalent rigid rod-like counterions: theory and simulations. J Phys A 42: 105401. doi: 10.1088/1751-8113/42/10/105401
|
| [31] | May S, Bohinc K (2014) Mean-field electrostatics of stiff rod-like ions, Eedited by: Dean D, Dobnikar J, Naji A and Podgornik R, lectrostatics of Soft and Disordered Matter, 1.st Eds., Pan Stanford Publishing, 335–346. |
| [32] |
Kim YW, Yi J, Pincus PA (2008) Attractions between Like-Charged Surfaces with DumbbellShaped Counterions. Phys Rev Lett 101: 208305. doi: 10.1103/PhysRevLett.101.208305
|
| [33] |
Grime MA, Khan MO, Bohinc K (2010) Interaction between Charged Surfaces Mediated by Rodlike Counterions: The Influence of Discrete Charge Distribution in the Solution and on the Surfaces. Langmuir 26: 6343–6349. doi: 10.1021/la903940j
|
| [34] |
Hatlo MM, Bohinc K, Lue L (2010) The properties of dimers confined between two charged plates. J Chem Phys 132: 114102. doi: 10.1063/1.3354121
|
| [35] |
Bohinc K, Reščič J, Maset S, et al. (2011) DebyeHckel theory for mixtures of rigid rodlike ions and salt. J Chem Phys 134: 074111-1-9. doi: 10.1063/1.3552226
|
| [36] |
Bohinc K, Grime JMA, Lue L (2012) The interactions between charged colloids with rod-like counterions. Soft matter 8: 5679–5686. doi: 10.1039/c2sm07463j
|
| [37] | Urbanija J, Bohinc K, Bellen A, et al. (2008) Attraction between negatively charged surfaces mediated by spherical counterions with quadrupolar charge distribution. J Chem Phys 129: 105101/1-5. |
| [38] |
Ibarra-Armenta JG, Mart´ ın-Molina A, Bohinc K, et al. (2012) Effects of the internal structure of spheroidal divalent ions on the charge density profiles of the electric double layer. J Chem Phys 137: 224701. doi: 10.1063/1.4768448
|
| [39] |
May S, Bohinc K (2011) Attraction between like charged surfaces mediated by uniformly charged spherical colloids in a salt solution. Croat Chem Acta 84: 251–257. doi: 10.5562/cca1824
|
| [40] |
Bohinc K, Reščič J, Dufreche JF, et al. (2013) Recycling of uranyl from contaminated water. J Phys Chem B 117: 10846–10851 251–257. doi: 10.1021/jp404822f
|
| [41] |
Gosule LC, Shellman JA (1976) Compact form of DNA induced by spermidine. Nature 259: 333–335. doi: 10.1038/259333a0
|
| [42] | Lerman LS (1971) A transition to a compact form of DNA in polymer solutions. PNAS USA 78: 1886–1890. |
| [43] | Jary D, Sikorav JL (1999) Cyclization of globular DNA. Implications for DNA-DNA interactions in vivo. Biochemistry 38: 3223–3227. |
| [44] | Vijayanathan V, Thomas T, Shirahata A, et al. (2001) DNA condensation by polyamines: a laser light scattering study of structural effects.Cyclization of globular DNA. Implications for DNADNA interactions in vivo. Biochemistry 40: 13644–13651. |
| [45] |
Slita AV, Kasyanenko NA, Nazarova OV, et al. (2007) DNA-polycation complexes: Effect of polycation structure on physico-chemical and biological properties. J Biotechnol 127: 679–693. doi: 10.1016/j.jbiotec.2006.07.016
|
| [46] | Slonitskii SV, Kuptsov V (1989) Binding of polyamines by the double-helical DNA molecule in unfolded and compact forms. Mol Biol (Mosk) 23: 507–517. |
| [47] |
Parsegian VA, Rand RP, Rau DC (2000) Osmotic stress, crowding, preferential hydration, and binding: A comparison of perspectives. Proc Natl Acad Sci U S A 97: 3987–3992. doi: 10.1073/pnas.97.8.3987
|
| [48] |
Strey HH, Podgornik R, Rau DC, et al. (1998) DNA-DNA interactions. Curr Opin Struct Biol 8: 309–313. doi: 10.1016/S0959-440X(98)80063-8
|
| [49] |
Marty R, N’soukpoe-Kossi CN, Charbonneau D, et al. (2009) Structural analysis of DNA complexation with cationic lipids. Nucl Acids Res 37: 849–857. doi: 10.1093/nar/gkn1003
|
| [50] | Hud NV, Vilfan ID (2005) Toroidal DNA condensates: unraveling the fine structure and the role of nucleation in determining size. Annu Rev Biophys Biomol Struct 34: 295–318. |
| [51] | Hansma HG, Kasuya K, Oroudjev E (2004) Atomic force microscopy imaging and pulling of nucleic acids. Curr Opin Struct Biol 14: 380. |
| [52] |
Keyser UF, van Dorp S, Lemay SG (2010) Tether forces in DNA electrophoresis. Chem Soc Rev 39: 939–47. doi: 10.1039/B902072C
|
| [53] |
Baumann CG, Bloomfield VA, Smith SB, et al. (2000) Stretching of single collapsed DNA molecules. Biophys J 78: 1965–1978. doi: 10.1016/S0006-3495(00)76744-0
|
| [54] |
Besteman K, Van Eijk K, Lemay SG (2007) Charge inversion accompanies DNA condensation by multivalent ions. Nat Phys 3: 641–644. doi: 10.1038/nphys697
|
| [55] |
Chien FT, Lin SG, Lai PY,et al. (2007) Observation of two forms of conformations in the reentrant condensation of DNA. Phys Rev E Stat Nonlin Soft Matter Phys 75: 041922. doi: 10.1103/PhysRevE.75.041922
|
| [56] |
Todd BA, Parsegian VA, Shirahata A, et al. (2008) Attractive forces between cation condensed DNA double helices. Biophys J 94: 4775–4782. doi: 10.1529/biophysj.107.127332
|
| [57] |
Bohinc K, Reščič J, Dufreche JF, et al. (2013) Recycling of Uranyl from Contaminated Water. J Phys Chem B 117: 10846–10851. doi: 10.1021/jp404822f
|
| [58] | Bohinc K, Lue L (2011) Interaction of similarly charged surfaces mediated by nanoparticles. Chin J Polymer Sci 29: 414–420. |
| [59] |
Grosberg AY, Nguyen TT, Shklovskii BI (2002) Colloquium: The physics of charge inversion in chemical and biological systems. Rev Mod Phys 74:329–345. doi: 10.1103/RevModPhys.74.329
|
| [60] |
Cherstvy AG, Teif V (2013) Structure-driven homology pairing of chromatin fibers: The role of electrostatics and protein bridging. J Biol Phys 39: 363–385. doi: 10.1007/s10867-012-9294-4
|
| [61] |
Cherstvy AG, Teif V (2014) Electrostatic effect of H1-histone protein binding on nucleosome repeat length. Phys Biol 11: 044001. doi: 10.1088/1478-3975/11/4/044001
|
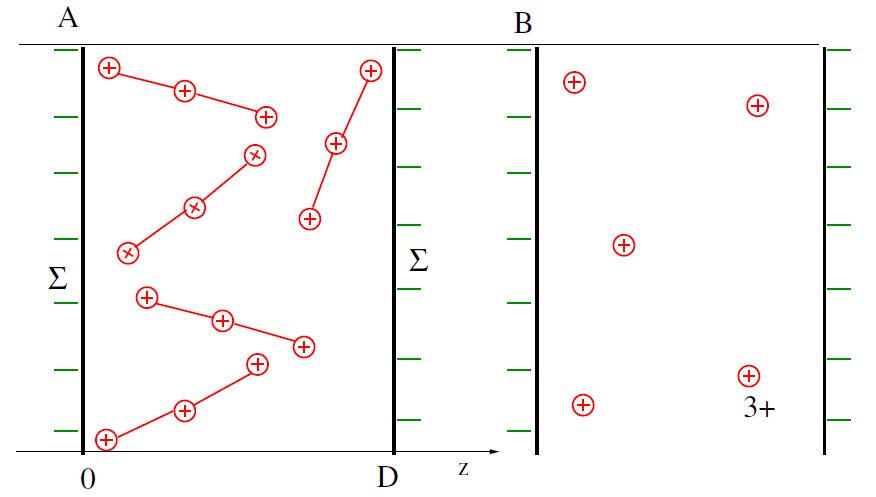
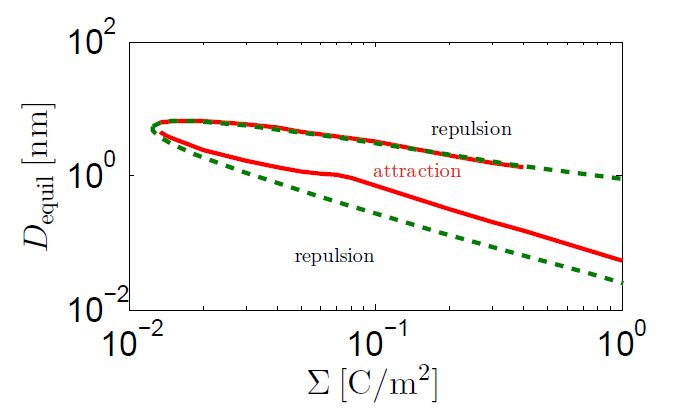
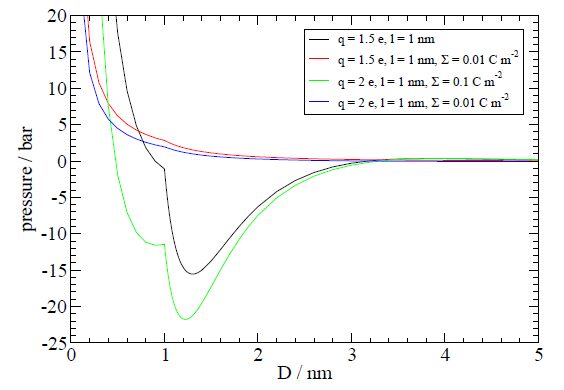
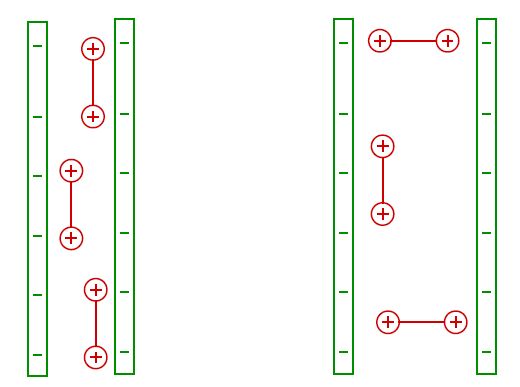
Figures(4)

Klemen Bohinc, Leo Lue. On the electrostatics of DNA in chromatin[J]. AIMS Biophysics, 2016, 3(1): 75-87. doi: 10.3934/biophy.2016.1.75







 DownLoad:
DownLoad: