Citation: Delores J. Grant, Leighcraft A. Shakes, Hope M. Wolf, Derek C. Norford, Pradeep K. Chatterjee. Exploring function of conserved non-coding DNA in its chromosomal context[J]. AIMS Biophysics, 2015, 2(4): 773-793. doi: 10.3934/biophy.2015.4.773
| [1] | Michael J. Hamilton, Matthew D. Young, Silvia Sauer, Ernest Martinez . The interplay of long non-coding RNAs and MYC in cancer. AIMS Biophysics, 2015, 2(4): 794-809. doi: 10.3934/biophy.2015.4.794 |
| [2] | Larisa A. Krasnobaeva, Ludmila V. Yakushevich . DNA kinks behavior in the potential pit-trap. AIMS Biophysics, 2022, 9(2): 130-146. doi: 10.3934/biophy.2022012 |
| [3] | Qiao Yi Chen, Max Costa, Hong Sun . Structure and function of histone acetyltransferase MOF. AIMS Biophysics, 2015, 2(4): 555-569. doi: 10.3934/biophy.2015.4.555 |
| [4] | Nick K. Olrichs, J. Bernd Helms . Novel insights into the function of the conserved domain of the CAP superfamily of proteins. AIMS Biophysics, 2016, 3(2): 232-246. doi: 10.3934/biophy.2016.2.232 |
| [5] | Larisa I. Fedoreyeva, Boris F. Vanyushin . Regulation of DNA methyltransferase gene expression by short peptides in nicotiana tabacum regenerants. AIMS Biophysics, 2021, 8(1): 66-79. doi: 10.3934/biophy.2021005 |
| [6] | Larisa I. Fedoreyeva, Boris F. Vanyushin, Ekaterina N. Baranova . Peptide AEDL alters chromatin conformation via histone binding. AIMS Biophysics, 2020, 7(1): 1-16. doi: 10.3934/biophy.2020001 |
| [7] | Angel Rivera-Calzada, Andrés López-Perrote, Roberto Melero, Jasminka Boskovic, Hugo Muñoz-Hernández, Fabrizio Martino, Oscar Llorca . Structure and Assembly of the PI3K-like Protein Kinases (PIKKs) Revealed by Electron Microscopy. AIMS Biophysics, 2015, 2(2): 36-57. doi: 10.3934/biophy.2015.2.36 |
| [8] | Shivakumar Keerthikumar . A catalogue of human secreted proteins and its implications. AIMS Biophysics, 2016, 3(4): 563-570. doi: 10.3934/biophy.2016.4.563 |
| [9] | Audrey Lejart, Gilles Salbert, Sébastien Huet . Cytosine hydroxymethylation by TET enzymes: From the control of gene expression to the regulation of DNA repair mechanisms, and back. AIMS Biophysics, 2018, 5(3): 182-193. doi: 10.3934/biophy.2018.3.182 |
| [10] | Andrea Bianchi, Chiara Lanzuolo . Into the chromatin world: Role of nuclear architecture in epigenome regulation. AIMS Biophysics, 2015, 2(4): 585-612. doi: 10.3934/biophy.2015.4.585 |
Cloaking using transformation optics (changes of variables) was introduced by Pendry, Schurig and Smith [30] for the Maxwell system and by Leonhardt [16] in the geometric optics setting. These authors used a singular change of variables, which blows up a point into a cloaked region. The same transformation had been used to establish (singular) non-uniqueness in Calderon's problem in [10]. To avoid using the singular structure, various regularized schemes have been proposed. One of them was suggested by Kohn, Shen, Vogelius and Weinstein [11], where instead of a point, a small ball of radius $ \varepsilon $ is blown up to the cloaked region. Approximate cloaking for acoustic waves has been studied in the quasistatic regime [11,26], the time harmonic regime [12,19,27,20], and the time regime [28,29], and approximate cloaking for electromagnetic waves has been studied in the time harmonic regime [4,14,24], see also the references therein. Finite energy solutions for the singular scheme have been studied extensively [9,32,33]. There are also other ways to achieve cloaking effects, such as the use of plasmonic coating [2], active exterior sources [31], complementary media [13,22], or via localized resonance [23] (see also [17,21]).
The goal of this paper is to investigate approximate cloaking for the heat equation using transformation optics. Thermal cloaking via transformation optics was initiated by Guenneau, Amra and Venante [8]. Craster, Guenneau, Hutridurga and Pavliotis [6] investigate the approximate cloaking for the heat equation using the approximate scheme in the spirit of [11]. They show that for the time large enough, the largeness depends on $ \varepsilon $, the degree of visibility is of the order $ \varepsilon^d $ ($ d = 2, 3 $) for sources that are independent of time. Their analysis is first based on the fact that as time goes to infinity, the solutions converge to the stationary states and then uses known results on approximate cloaking in the quasistatic regime [11,26].
In this paper, we show that approximate cloaking is achieved at any positive time and established the degree of invisibility of order $ \varepsilon $ in three dimensions and $ |\ln \varepsilon|^{-1} $ in two dimensions. Our results hold for a general source that depends on both time and space variables, and our estimates depend only on the range of the materials inside the cloaked region. The degree of visibility obtained herein is optimal due to the fact that a finite time interval is considered (compare with [6]). The analysis in this paper is of frequency type via Fourier transform with respect to time. This approach is robust and can be used in different context. A technical issue is on the blow up of the fundamental solution of the Helmholtz type equations in two dimensions in the low frequency regime. We emphasize that even though our setting is in a bounded domain, we employs Fourier transform in time instead of eigenmodes decomposition. This has the advantage that one can put the non-perturbed system and the cloaking system in the same context.
We next describe the problem in more detail and state the main result. Our starting point is the regularization scheme [11] in which a transformation blows up a small ball $ B_\varepsilon $ ($ 0 < \varepsilon < 1/2 $) instead of a point into the cloaked region $ B_1 $ in $ \mathbb{R}^d $ ($ d = 2, 3 $). Here and in what follows, for $ r > 0 $, $ B_r $ denotes the ball centered at the origin and of radius $ r $ in $ \mathbb{R}^d $. Our assumption on the geometry of the cloaked region is mainly to simplify the notations. Concerning the transformation, we consider the map $ F_{\varepsilon}: {\mathbb{R }}^d \rightarrow {\mathbb{R }}^d $ defined by
| $ Fε(x)={x in Rd∖B2,(2−2ε2−ε+|x|2−ε)x|x| in B2∖Bε,xε in Bε. $ | (1.1) |
In what follows, we use the standard notations
| $ F∗A(y)=∇F(x)A(x)∇FT(x)|det∇F(x)|,F∗ρ(y)=ρ(x)|det∇F(x)|,x=F−1(y), $ | (1.2) |
for the "pushforward" of a symmetric, matrix-valued function $ A $, and a scalar function $ \rho $, by the diffeomorphism $ F $, and $ I $ denotes the identity matrix. The cloaking device in the region $ B_2 \setminus B_1 $ constructed from the transformation technique is given by
| $ (Fε∗I,Fε∗1) in B2∖B1, $ | (1.3) |
a pair of a matrix-valued function and a function that characterize the material properties in $ B_2 \setminus B_1 $. Physically, this is the pair of the thermal diffusivity and the mass density of the material.
Let $ \Omega $ with $ B_2 \Subset \Omega \subset \mathbb{R}^d $ ($ d = 2, 3 $)* be a bounded region for which the heat flow is considered. Suppose that the medium outside $ B_2 $ (the cloaking device and the cloaked region) is homogeneous so that it is characterized by the pair $ (I, 1) $, and the cloaked region $ B_1 $ is characterized by a pair $ (a_O, \rho_O) $ where $ a_O $ is a matrix-valued function and $ \rho_O $ is a real function, both defined in $ B_{1} $. The medium in $ \Omega $ is then given by
*The notation $ D \Subset \Omega $ means that the closure of $ D $ is a subset of $ \Omega $.
| $ (Ac,ρc)={(I,1) in Ω∖B2,(Fε∗I,Fε∗1) in B2∖B1,(aO,ρO) in B1. $ | (1.4) |
In what follows, we make the usual assumption that $ a_O $ is symmetric and uniformly elliptic and $ \rho_O $ is a positive function bounded above and below by positive constants, i.e., for a.e. $ x \in B_{1} $,
| $ Λ−1|ξ|2≤⟨aO(x)ξ,ξ⟩≤Λ|ξ|2 for all ξ∈Rd, $ | (1.5) |
and
| $ Λ−1≤ρO(x)≤Λ, $ | (1.6) |
for some $ \Lambda \ge 1 $. Given a function $ f \in L^1 \big((0, + \infty), L^2(\Omega) \big) $ and an initial condition $ u_0 \in L^2(\Omega) $, in the medium characterzied by $ (A_c, \rho_c) $, one obtains a unique weak solution $ u_c \in L^2\big((0, \infty); H^1(\Omega)\big) $ $ \cap C \big([0, + \infty); L^2(\Omega) \big) $ of the system
| $ {∂t(ρcuc)−div(Ac∇uc)=f in (0,+∞)×Ω,uc=0 on (0,+∞)×∂Ω,uc(t=0,⋅)=u0 in Ω, $ | (1.7) |
and in the homogeneneous medium characterized by $ (I, 1) $, one gets a unique weak solution $ u \in L^2\big((0, \infty); H^1(\Omega) \big) \cap C \big([0, + \infty); L^2(\Omega) \big) $ of the system
| $ {∂tu−Δu=f in (0,+∞)×Ω,uc=0 on (0,+∞)×∂Ω,uc(t=0,⋅)=u0 in Ω. $ | (1.8) |
The approximate cloaking meaning of the scheme (1.4) is given in the following result:
Theorem 1.1. Let $ u_0 \in L^2(\Omega) $ and $ f \in L^1\big((0, + \infty); L^2(\Omega) \big) $ be such that $ \operatorname{supp} u_0, \, \operatorname{supp} f(t, \cdot) \subset \Omega \setminus B_2 $ for $ t > 0 $. Assume that $ u_c $ and $ u $ are the solution of (1.7) and (1.8) respectively. Then, for $ 0 < \varepsilon < 1/2 $,
| $ ‖uc(t,⋅)−u(t,⋅)‖H1(Ω∖B2)≤Ce(ε,d)(‖f‖L1((0,+∞);L2(Ω))+‖u0‖L2(Ω)), $ |
for some positive constant $ C $ depending on $ \Lambda $ but independent of $ f $, $ u_0 $, and $ \varepsilon $, where
| $ e(\varepsilon, d) = \left\{ εif d=3,|lnε|−1if d=2.\right. $ |
As a consequence of Theorem 1.1, $ \lim_{\varepsilon \to 0} u_c (t, \cdot) = u (t, \cdot) $ in $ (0, + \infty) \times (\Omega \setminus B_2) $ for all $ f $ with compact support outside $ (0, +\infty) \times B_2 $ and for all $ u_0 $ with compact support outside $ B_2 $. One therefore cannot detect the difference between $ (A_c, \rho_c) $ and $ (I, 1) $ as $ \varepsilon \rightarrow 0 $ by observation of $ u_c $ outside $ B_2 $: Cloaking is achieved for observers outside $ B_2 $ in the limit as $ \varepsilon \rightarrow 0 $.
We now briefly describe the idea of the proof. The starting point of the analysis is the invariance of the heat equations under a change of variables which we now state.
Lemma 1.1. Let $ d \ge 2 $, $ T > 0 $, $ \Omega $ be a bounded open subset of $ \mathbb{R}^d $ of class $ C^1 $, and let $ A $ be an elliptic symmetric matrix-valued function, and $ \rho $ be a bounded, measurable function defined on $ \Omega $ bounded above and below by positive constants. Let $ F: \Omega \mapsto \Omega $ be bijective such that $ F $ and $ F^{-1} $ are Lipschitz, $ \det \nabla F > c $ for a.e. $ x \in \Omega $ for some $ c > 0 $, and $ F(x) = x $ near $ \partial \Omega $. Let $ f \in L^1\big((0, T); L^2(\Omega) \big) $ and $ u_0 \in L^2(\Omega) $. Then $ u \in L^2\big((0, T); H^1_0(\Omega) \big) \cap C\big([0, T); L^2(\Omega) \big) $ is the weak solution of
| $ {∂t(ρu)−div(A∇u)=fin ΩT,u=0on (0,T)×∂Ω,u(0,⋅)=u0in Ω, $ | (1.9) |
if and only if $ v(t, \cdot): = u (t, \cdot) \circ F^{-1} \in L^2\big((0, T); H^1_0(\Omega) \big) \cap C\big([0, T); L^2(\Omega) \big) $ is the weak solution of
| $ {∂t(F∗ρv)−div(F∗A∇v)=F∗fin ΩT,u=0on (0,T)×∂Ω,v(0,⋅)=u0∘F−1in Ω. $ | (1.10) |
Recall that $ F_* $ is defined in (1.2). In this paper, we use the following standard definition of weak solutions:
Definition 1.1. Let $ d \ge 2 $ and $ T > 0 $. We say a function
| $ u∈L2((0,T);H10(Ω))∩C([0,T);L2(Ω)) $ |
is a weak solution to (1.9) if $ u(0, \cdot) = u_0 \mbox{ in } \Omega $ and $ u $ satisfies
| $ ddt∫Ωρu(t,⋅)φ+∫ΩA∇u(t,⋅)∇φ=∫Ωf(t,⋅)φ in (0,T), $ | (1.11) |
in the distributional sense for all $ \varphi \in H^1_0(\Omega) $.
The existence and uniqueness of weak solutions are standard, see, e.g., [1] (in fact, in [1], $ f $ is assumed in $ L^2\big((0, T); L^2(\Omega) \big) $, however, the conclusion holds also for $ f \in L^1\big((0, T); L^2(\Omega) \big) $ with a similar proof, see, e.g., [25]). The proof of Lemma 1.1 is similar to that of the Helmholtz equation, see, e.g., [12] (see also [6] for a parabolic version).
We now return to the idea of the proof of Theorem 1.1. Set
| $ u_\varepsilon (t, \cdot) = u_c (t, \cdot) \circ F_\varepsilon^{-1} \mbox{ for } t \in (0, +\infty). $ |
Then $ u_\varepsilon $ is the unique solution of the system
| $ {∂t(ρεuε)−div(Aε∇uε)=f in (0,+∞)×Ω,uε=0 on (0,+∞)×∂Ω,uε(t=0,⋅)=u0 in Ω, $ | (1.12) |
where
| $ (Aε,ρε)={(I,1) in Ω∖Bε,(ε2−daO(⋅/ε),ε−dρO(⋅/ε)) in Bε. $ | (1.13) |
Moreover,
| $ u_c - u = u_\varepsilon - u \mbox{ in } (0, + \infty) \times (\Omega \setminus B_2). $ |
In comparing the coefficients of the systems verified by $ u $ and $ u_\varepsilon $, the analysis can be derived from the study of the effect of a small inclusion $ B_\varepsilon $. The case in which finite isotropic materials contain inside the small inclusion was investigated in [3] (see also [5] for a related context). The analysis in [3] partly involved the polarization tensor information and took the advantage of the fact that the coefficients inside the small inclusion are finite. In the cloaking context, Craster et al. [6] derived an estimate of the order $ \varepsilon^d $ for a time larger than a threshold one. Their analysis is based on long time behavior of solutions to parabolic equations and estimates for the degree of visibility of the conducting problem, see [11,26], hence the threshold time goes to infinity as $ \varepsilon \to 0 $.
In this paper, to overcome the blow up of the coefficients inside the small inclusion and to achieve the cloaking effect at any positive time, we follow the approach of Nguyen and Vogelius in [28]. The idea is to derive appropriate estimates for the effect of small inclusions in the time domain from the ones in the frequency domain using the Fourier transform with respect to time. Due to the dissipative nature of the heat equation, the problem in the frequency for the heat equation is more stable than the one corresponding to the acoustic waves, see, e.g., [27,28], and the analysis is somehow easier to handle in the high frequency regime. After using a standard blow-up argument, a technical point in the analysis is to obtain an estimate for the solutions of the equation $ \Delta v + i \omega \varepsilon^2 v = 0 $ in $ \mathbb{R}^d \setminus B_1 $ ($ \omega > 0 $) at the distance of the order $ 1/\varepsilon $ in which the dependence on $ \varepsilon $ and $ \omega $ are explicit (see Lemma 2.2). Due to the blow up of the fundamental solution in two dimensions, the analysis requires new ideas. We emphasize that even though our setting is in a bounded domain with zero Dirichlet boundary condition, we employs Fourier transform in time instead of eigenmodes decomposition as in [6] to put both systems of $ u_\varepsilon $ and $ u $ in the same context.
To implement the analysis in the frequency domain, let us introduce the Fourier transform with respect to time $ t $:
| $ ˆφ(k,x)=∫Rφ(t,x)eiktdt for k∈R, $ | (2.1) |
for $ \varphi \in L^2((-\infty, + \infty), L^2(\mathbb{R}^d)) $. Extending $ u, \, u_c $, $ u_\rho $ and $ f $ by $ 0 $ for $ t < 0 $, and considering the Fourier with respect to time at the frequency $ \omega > 0 $, we obtain
| $ Δˆu+iωˆu=−(ˆf+u0) in Ω, $ |
and
| $ div(Aε∇ˆuε)+iωρεˆuε=−(ˆf+u0) in Ω, $ |
where
| $ (Aε,ρε)={(I,1) in Ω∖Bε,(ε2−daO(⋅/ε),ε−dρO(⋅/ε)) in Bε. $ |
The main ingredient in the proof of Theorem 1.1 is the following:
Proposition 2.1. Let $ \omega > 0 $, $ 0 < \varepsilon < 1/2 $, and let $ g \in L^2(\Omega) $ with $ \operatorname{supp} g \subset \Omega \setminus B_2 $. Assume that $ v, \, v_\varepsilon \in H^1(\Omega) $ are respectively the unique solution of the systems
| $ {Δv+iωv=gin Ω,v=0on ∂Ω, $ |
and
| $ {div(Aε∇vε)+iωρεvε=gin Ω,vε=0on ∂Ω. $ |
We have
| $ ‖vε−v‖H1(Ω∖B2)≤Ce(ε,ω,d)(1+ω−1/2)‖g‖L2(Ω), $ | (2.2) |
for some positive constant $ C $ independent of $ \varepsilon $, $ \omega $ and $ g $. Here
| $ e(ε,ω,3)=εe−ω1/2/4, $ | (2.3) |
and
| $ e(ε,ω,2)={e−ω1/2/4/|lnε|if ω≥1/2,lnω/ln(ωε)if 0<ω<1/2. $ | (2.4) |
The rest of this section is divided into three subsections. In the first subsection, we present several lemmas used in the proof of Proposition 2.1. The proofs of Proposition 2.1 and Theorem 1.1 are then given in the second and the third subsections, respectively.
In this subsection, we state and prove several useful lemmas used in the proof of Proposition 2.1. Throughout, $ D \subset B_1 $ denotes a smooth, bounded, open subset of $ \mathbb{R}^d $ such that $ \mathbb{R}^d \setminus D $ is connected, and $ \nu $ denotes the unit normal vector field on $ \partial D $, directed into $ \mathbb{R}^d \setminus D $.
The first result is the following simple one:
Lemma 2.1. Let $ d = 2, \, 3 $, $ k > 0 $, and let $ v \in H^1 ({\mathbb{R }}^d \setminus D) $ be such that $ \Delta v +i k v = 0 $ in $ \mathbb{R}^d \setminus D $. We have, for $ R > 2 $,
| $ ‖v‖H1(BR∖D)≤CR(1+k)‖v‖H1/2(∂D), $ | (2.5) |
for some positive constants $ C_R $ independent of $ k $ and $ v $.
Proof. Multiplying the equation by $ \bar v $ (the conjugate of $ v $) and integrating by parts, we have
| $ \int_{ \mathbb{R}^d \setminus D} |\nabla v|^2 - i k \int_{ \mathbb{R}^d \setminus D} |v|^2 = \int_{\partial D} \partial_\nu v \bar v. $ |
This implies
| $ ∫Rd∖D|∇v|2+k∫Rd∖D|v|2≤C‖∂νv‖H−1/2(∂D)‖v‖H1/2(∂D). $ | (2.6) |
Here and in what follows, $ C $ denotes a positive constant independent of $ v $ and $ k $. Since $ \Delta v = - i k v $ in $ B_2 \setminus D $, by the trace theory, see, e.g., [7,Theorem 2.5], we have
| $ ‖∂νv‖H−1/2(∂D)≤C(‖∇v‖L2(B2∖D)+‖Δv‖L2(B2∖D))≤C(‖∇v‖L2(B2∖D)+k‖v‖L2(B2∖D)). $ | (2.7) |
Combining (2.6) and (2.7) yields
| $ ∫Rd∖D|∇v|2+k∫Rd∖D|v|2≤C(1+k)‖v‖2H1/2(∂D). $ | (2.8) |
The conclusion follows when $ k \ge 1 $.
Next, consider the case $ 0 < k < 1 $. In the case where $ d = 3 $, the conclusion is a direct consequence of (2.8) and the Hardy inequality (see, e.g., [18,Lemma 2.5.7]):
| $ ∫R3∖D|v|2|x|2≤C∫R3∖D|∇v|2. $ | (2.9) |
We next consider the case where $ d = 2 $. One just needs to show
| $ ∫BR∖D|v|2≤C‖v‖2H1/2(∂D). $ | (2.10) |
By the Hardy inequality (see, e.g., [18,Lemma 2.5.7]),
| $ ∫R2∖D|v|2|x|2ln(2+|x|)2≤C(∫R2∖D|∇v|2+∫B2∖D|v|2), $ | (2.11) |
it suffices to prove (2.10) for $ R = 2 $ by contradiction. Suppose that there exists a sequence $ (k_n) \to 0 $ and a sequence $ (v_n) \in H^1(\mathbb{R}^2 \setminus D) $ such that
| $ Δvn+iknvn=0 in R2∖D,‖vn‖L2(B2∖D)=1, and limn→+∞‖vn‖H1/2(∂D)=0. $ |
Denote
| $ W1(R2∖D)={u∈L1loc(R2∖D);u(x)ln(2+|x|)√1+|x|2∈L2(R2∖D) and ∇u∈L2(R2∖D)}. $ |
By (2.8) and (2.11), one might assume that $ v_n $ converges to $ v $ weakly in $ H^1_{ \operatorname{loc }}(\mathbb{R}^2 \setminus D) $ and strongly in $ L^2(B_2 \setminus D) $. Moreover, $ v \in W^1(\mathbb{R}^2 \setminus D) $ and $ v $ satisfies
| $ Δv=0 in R2∖D,v=0 on ∂D, $ | (2.12) |
and
| $ ‖v‖L2(B2∖D)=1. $ | (2.13) |
From (2.12), we have $ v = 0 $ in $ \mathbb{R}^2 \setminus D $ (see, e.g., [18]) which contradicts (2.13). The proof is complete.
We also have
Lemma 2.2. Let $ d = 2, 3 $, $ \omega > 0 $, $ 0 < \varepsilon < 1/2 $, and let $ v \in H^1 ({\mathbb{R }}^d \setminus D) $ be a solution of $ \Delta v +i\omega \varepsilon^2 v = 0 $ in $ \mathbb{R}^d \setminus D $. We have, for $ 3/2 < |x| < R $,
| $ |v(x/ε)|≤Ce(ε,ω,d)‖v‖H1/2(∂D), $ | (2.14) |
for some positive constant $ C = C_{R} $ independent of $ \varepsilon $, $ \omega $ and $ v $.
Recall that $ e(\varepsilon, \omega, d) $ is given in (2.3) and (2.4).
Proof. By the trace theory and the regularity theory of elliptic equations, we have
| $ ‖v‖L2(∂B2)+‖∇v‖L2(∂B2)≤C‖v‖H2(B5/2∖B3/2)≤C(1+ω1/2ε)‖v‖H1(B3∖B1). $ | (2.15) |
It follows from Lemma 2.1 that
| $ ‖v‖L2(∂B2)+‖∇v‖L2(∂B2)≤C(1+ω3/2)‖v‖H1/2(∂D). $ | (2.16) |
Here and in what follows in this proof, $ C $ denotes a positive constant depending only on $ R $ and $ D $.
The representation formula gives
| $ v(x)=∫∂B2(Gℓ(x,y)∂rv(y)−∂ryGℓ(x,y)v(y))dy for x∈Rd∖ˉB2, $ | (2.17) |
where $ \ell = e^{i \pi/4} \varepsilon \omega^{1/2} $, and, for $ x \neq y $,
| $ G_\ell(x, y) = \frac{e^{i\ell |x-y| }}{4 \pi |x -y|} \mbox{ if } d = 3 \quad \mbox{ and } \quad G_\ell(x, y) = \frac{i}{4}H^{(1)}_0 (\ell|x-y|) \mbox{ if } d = 2. $ |
Here $ H^{(1)}_0 $ is the Hankel function of the first kind of order 0. Recall, see, e.g., [15,Chapter 5], that
| $ H(1)0(z)=2iπln|z|2+1+2iγπ+O(|z|2log|z|) as z→0,z∉(−∞,0], $ | (2.18) |
and
| $ H(1)0(z)=√2πzei(z+π4)(1+O(|z|−1))z→∞,z∉(−∞,0]. $ | (2.19) |
We now consider the case $ d = 3 $. We have, for $ 3/2 < |x| < R $ and $ y \in \partial B_{2} $,
| $ |e^{i \ell |x/ \varepsilon - y|}| \le e^{- \frac{\sqrt{2 }}{2} \omega^{1/2} |x - \varepsilon y|} \le e^{- \omega^{1/2} |x|/ 3}. $ |
It follows that, for $ 3/2 < |x| < R $ and $ y \in \partial B_{2} $,
| $ |Gℓ(x/ε,y)|≤Cεe−3ω1/2/10. $ | (2.20) |
Similarly, one has, for $ 3/2 < |x| < R $ and $ y \in \partial B_{2} $,
| $ |∂ryGℓ(x/ε,y)|≤C(ε2ω1/2|x|+ε2|x|2)e−ω1/2|x|/3≤Cεe−3ω1/2/10. $ | (2.21) |
Combining (2.17), (2.20) and (2.21) yields
| $ |v(x/ε)|≤Cεe−3ω1/2/10(‖v‖L2(∂B2)+‖∇v‖L2(∂B2)) for 3/2<|x|<R. $ |
We derive from (2.16) that
| $ |v(x/ε)|≤Cεe−ω1/2/4‖v‖H1/2(∂D) for 3/2<|x|<R; $ |
which is the conclusion in the case $ d = 3 $.
We next deal with the case where $ d = 2 $ and $ \omega > \varepsilon^{-2}/4 $, which is equivalent to $ |\ell| > 1/2 $. From (2.19), we derive that, for $ 3/2 < |x| < R $ and $ y \in \partial B_{2} $,
| $ |Gℓ(x/ε,y)|≤Cω−1/4e−3ω1/2/10 and |∂ryGℓ(x/ε,y)|≤Cεω1/4e−3ω1/2/10. $ | (2.22) |
Using (2.16) and combining (2.17) and (2.22), we obtain, since $ \omega > \varepsilon^{-2}/4 $,
| $ |v(x/ε)|≤Cεe−ω1/2/4‖v‖H1/2(∂D) for 3/2<|x|<R, $ |
which gives the conclusion in this case.
We finally deal with the case where $ d = 2 $ and $ 0 < \omega < \varepsilon^{-2}/4 $, which is equivalent to $ |\ell| < 1/2 $. From (2.17), we obtain, for $ x \in \partial B_{4} $,
| $ v(x)=∫∂B2([Gℓ(x,y)−Gℓ(x,0)]∂rv(y)−∂ryGℓ(x,y)v(y))dy+∫∂B2Gℓ(x,0)∂rv(y)dy. $ | (2.23) |
Since $ d = 2 $, we have
| $ ‖v‖L∞(B5∖B3)≤C‖v‖H2(B5∖B3)≤C‖v‖H2(B5∖B2)≤C(1+ω1/2)‖v‖H1(B6∖B1). $ |
It follows from Lemma 2.1 and the trace theory that
| $ ‖v‖L∞(B5∖B3)+‖v‖L2(∂B2)+‖∇v‖L2(∂B2)≤C(1+ω3/2)‖v‖H1/2(∂D). $ | (2.24) |
Since, by (2.18),
| $ |\nabla_y G_\ell(x, y)| \le C \mbox{ for } x \in \partial B_{4} \mbox{ and } y \in \partial B_{2} $ |
and
| $ |G_\ell(x, 0)|\ge C|\ln|\ell|| \mbox{ for } x \in \partial B_4, $ |
we derive from (2.23) and (2.24) that
| $ |∫∂B2∂rv(y)dy|≤C(1+ω3/2)|ln|ℓ||‖v‖H1/2(∂D). $ | (2.25) |
Again using (2.17), we get, for $ 3/2 < |x| < R $,
| $ v(x/ε)=∫∂B2([Gℓ(x/ε,y)−Gℓ(x/ε,0)]∂rv(y)−∂ryGℓ(x/ε,y)v(y))dy+∫∂B2Gℓ(x/ε,0)∂rv(y)dy. $ | (2.26) |
Since, by (2.18), for $ 0 < \omega < 1/2 $,
| $ |G_{\ell} (x/ \varepsilon, 0)| \le C |\ln \omega| \quad \mbox{ and } \quad |\nabla_y G_\ell(x/\varepsilon, y)| \le C \varepsilon \quad \mbox{ for } 3/2 \lt |x| \lt R, \, y \in \partial B_{2}, $ |
and, by (2.19), for $ 1/2 < \omega < \varepsilon^{-2}/4 $,
| $ |G_\ell(x/\varepsilon, 0)| \le C \omega^{-1/4} e^{- 3\omega^{1/2}/10} \mbox{ and } |\nabla_y G_\ell(x/\varepsilon, y)| \le C\varepsilon \omega^{1/4} e^{- 3\omega^{1/2}/ 10} \mbox{ for } 3/2 \lt |x| \lt R, \, y \in \partial B_{2}, $ |
we derive from (2.24), (2.25) and (2.26) that, for $ 3/2 < |x| < R $,
| $ \begin{equation*} |v(x/ \varepsilon)| \le \left\{ \begin{array}{cl} \frac{C|\ln \omega| }{|\ln |\ell||} \|v \| _{H^{1/2}(\partial D)} & \mbox{ if } 0 \lt \omega \lt 1/2, \\[6pt] \frac{C \omega^{3/2} e^{- 3\omega^{1/2}/10 }}{|\ln |\ell||} \|v \| _{H^{1/2}(\partial D)} & \mbox{ if } 1/2 \lt \omega \lt \varepsilon^{-2}/4, \end{array} \right. \end{equation*} $ |
which yields the conclusion in the case $ 0 < \omega < \varepsilon^{-2}/4 $. The proof is complete.
In this proof, $ C $ denotes a positive constant depending only on $ \Omega $ and $ \Lambda $. Multiplying the equation of $ v_\varepsilon $ by $ \bar v_\varepsilon $ and integrating in $ \Omega $, we derive that
| $ \begin{equation} \int_{\Omega} \langle A_\varepsilon \nabla v_\varepsilon, \nabla v_\varepsilon \rangle + \omega \int_{\Omega} \rho_\varepsilon |v_\varepsilon|^2 \le C \| g\|_{L^2(\Omega)}^2. \end{equation} $ | (2.27) |
Here we used Poincaré's inequality
| $ \| v_\varepsilon \|_{L^2(\Omega)} \le C \| \nabla v_\varepsilon\|_{L^2(\Omega)}. $ |
It follows from (2.27) that
| $ \begin{align} \| v_{\varepsilon} (\varepsilon \, \cdot \, ) \|_{H^{1/2}(\partial B_1)}^2 &\le C \| v_{\varepsilon} (\varepsilon \, \cdot \, ) \|_{H^{1}(B_1)}^2 \\[6pt] &\le C \int_{B_\varepsilon} \frac{1}{\varepsilon^{d-2 }}|\nabla v_\varepsilon|^2 + \frac{1}{\varepsilon^d} |v_\varepsilon|^2 \le C (1+\omega^{-1}) \| g\|_{L^2(\Omega)}^2. \end{align} $ | (2.28) |
Similarly, using the equation for $ v $ and Poincaré's inequality, we get
| $ \begin{equation} \|v\|_{H^1(\Omega)} \le C \| g\|_{L^2(\Omega)}. \end{equation} $ | (2.29) |
Since $ \Delta v+i\omega v = 0 $ in $ B_2 $, using Caccioppolli's inequality, we have
| $ \begin{equation} \|v\|_{H^3(B_{1})} \le C\|v\|_{H^2(B_{3/2})} \le C\|v\|_{H^1(B_{2})}\le C \| g\|_{L^2(\Omega)}. \end{equation} $ | (2.30) |
By Sobolev embedding, as $ d\le 3 $,
| $ \begin{equation} \| v\|_{W^{1, \infty}(B_1)} \le C \|v\|_{H^3(B_{1})} . \end{equation} $ | (2.31) |
It follows that
| $ \begin{equation} \| v(\varepsilon \, \cdot \, ) \|_{H^{1/2}(\partial B_1)} \le C \| v(\varepsilon \, \cdot \, )\|_{H^1(B_1)} \le C \| v\|_{W^{1, \infty}(B_1)} \le C \| g\|_{L^2(\Omega)}. \end{equation} $ | (2.32) |
Set
| $ w_{\varepsilon} = v_{\varepsilon} - v \mbox{ in } \Omega \setminus B_\varepsilon. $ |
Then $ w_{\varepsilon} \in H^1(\Omega \setminus B_\varepsilon) $ and satisfies
| $ \begin{equation} \left\{ \begin{array}{cl} \Delta w_{\varepsilon} + i \omega w_{\varepsilon} = 0 &\mbox{ in } \Omega \setminus B_\varepsilon, \\[6pt] w_{\varepsilon} = v_\varepsilon - v &\mbox{ on } \partial B_{\varepsilon}, \\[6pt] w_{\varepsilon} = 0 &\mbox{ on } \partial \Omega. \end{array}\right. \end{equation} $ | (2.33) |
Let $ \widetilde w_{\varepsilon}\in H^1(\mathbb{R}^d \setminus B_{\varepsilon}) $ be the unique solution of the system
| $ \begin{equation} \left\{ \begin{array}{cl} \Delta \widetilde w_{\varepsilon} + i \omega \widetilde w_{\varepsilon} = 0 & \mbox{ in } \mathbb{R}^d \setminus B_{\varepsilon}, \\[6pt] \widetilde w_{\varepsilon} = w_\varepsilon & \mbox{ on } \partial B_{\varepsilon}, \end{array}\right. \end{equation} $ | (2.34) |
and set
| $ \widetilde W_{\varepsilon} = \widetilde w_{\varepsilon}(\varepsilon \, \cdot \, ) \mbox{ in } \mathbb{R}^d \setminus B_1. $ |
Then $ \widetilde W_{\varepsilon} \in H^1(\mathbb{R}^d \setminus B_{1}) $ is the unique solution of the system
| $ \begin{equation} \left\{ \begin{array}{cl} \Delta \widetilde W_{\varepsilon} + i \omega \varepsilon^2 \widetilde W_{\varepsilon} = 0 & \mbox{ in } \mathbb{R}^d \setminus B_{1}, \\[6pt] \widetilde W_{\varepsilon} = w_\varepsilon (\varepsilon \, \cdot \, ) & \mbox{ on } \partial B_{1}. \end{array}\right. \end{equation} $ | (2.35) |
Fix $ r_0 > 2 $ such that $ \Omega \subset B_{r_0} $. By Lemma 2.2, we have, for $ 1 \le |x| < r_0 $, that
| $ \begin{equation*} | \widetilde W_{\varepsilon} (x / \varepsilon)| \le C e(\varepsilon, \omega, d) \|w_\varepsilon (\varepsilon \, \cdot \, ) \|_{H^{1/2}(\partial B_1)}, \end{equation*} $ |
which yields, for $ x \in B_{r_0} \setminus B_{1} $, that
| $ \begin{equation*} | \widetilde w_{\varepsilon}(x)| \le C e(\varepsilon, \omega, d) \|w_\varepsilon (\varepsilon \, \cdot \, ) \|_{H^{1/2}(\partial B_1)}. \end{equation*} $ |
Since $ \Delta \widetilde w_{\varepsilon} + i \omega \widetilde w_{\varepsilon} = 0 $ in $ B_{r_0} \setminus B_{1} $, it follows from Caccioppoli's inequality that
| $ \begin{equation} \| \widetilde w_{\varepsilon} \|_{H^1(B_2 \setminus B_{3/2})} \le C e(\varepsilon, \omega, d) \|w_\varepsilon (\varepsilon \, \cdot \, ) \|_{H^{1/2}(\partial B_1)}. \end{equation} $ | (2.36) |
Fix $ \varphi \in C^2(\mathbb{R}^d) $ such that $ \varphi = 1 $ in $ B_{3/2} $ and $ \varphi = 0 $ in $ \mathbb{R}^d \setminus B_2 $, and set
| $ \chi_{\varepsilon} = w_{\varepsilon} - \varphi \widetilde w_{\varepsilon} \mbox{ in } \Omega \setminus B_\varepsilon. $ |
Then $ \chi_{\varepsilon}\in H^1_0(\Omega \setminus B_\varepsilon) $ and satisfies
| $ \begin{equation*} \Delta \chi_{\varepsilon} +i\omega \chi_{\varepsilon} = -\Delta \varphi \widetilde w_{\varepsilon} - 2\nabla \varphi \cdot \nabla \widetilde w_{\varepsilon} \mbox{ in } \Omega\setminus B_\varepsilon. \end{equation*} $ |
Multiplying the equation of $ \chi_{\varepsilon} $ by $ \bar \chi_{\varepsilon} $ and integrating by parts, we obtain, by Poincaré's inequality,
| $ \begin{equation} \| \chi_{\varepsilon} \|_{H^1(\Omega \setminus B_\varepsilon)}\le C \| \widetilde w_{\varepsilon} \|_{H^1(B_2 \setminus B_{3/2})}. \end{equation} $ | (2.37) |
Combining (2.36) and (2.37) yields
| $ \begin{equation} \| w_{\varepsilon} \|_{H^1(\Omega \setminus B_2)} \le C e(\varepsilon, \omega, d) \|w_\varepsilon (\varepsilon \, \cdot \, ) \|_{H^{1/2}(\partial B_1)}. \end{equation} $ | (2.38) |
The conclusion now follows from (2.28) and (2.32).
Let $ v_\varepsilon = u_\varepsilon - u $. Using the fact that $ v_\varepsilon $ is real, by the inversion theorem and Minkowski's inequality, we have, for $ t > 0 $,
| $ \begin{align} \|v_\varepsilon(t, \cdot) \|_{L^2(\Omega \setminus B_2)} \le C \int_0^\infty \|\hat{v}_\varepsilon (\omega, \cdot) \|_{L^2(\Omega \setminus B_2) } \, d \omega. \end{align} $ | (2.39) |
Using Proposition 2.1, we get
| $ \begin{align*} \int_0^\infty \|\hat{v}_\varepsilon (\omega, \cdot) \|_{L^2(\Omega \setminus B_2) } \, d \omega &\le C \int_0^\infty (1+\omega^{-1/2})e(\varepsilon, \omega, d)\| \hat{f}(\omega)+u_0\|_{L^2(\Omega \setminus B_2) } \, d \omega \\[6pt] & \le C \mbox{esssup}_{\omega \gt 0} \|\hat{f}(\omega)+u_0\|_{L^2(\Omega \setminus B_2) } \int_0^\infty (1+\omega^{-1/2})e(\varepsilon, \omega, d) \, d \omega \\[6pt] & \le Ce(\varepsilon, d) \big(\|f\|_{L^1\big( (0, + \infty); L^2(\Omega) \big)} +\|u_0\|_{L^2(\Omega)} \big). \end{align*} $ |
It follows from (2.39) that, for $ t > 0 $,
| $ \begin{align*} \|v_\varepsilon(t, \cdot) \|_{L^2(\Omega \setminus B_2)} \le Ce(\varepsilon, d) \big(\|f\|_{L^1\big( (0, + \infty); L^2(\Omega) \big)} +\|u_0\|_{L^2(\Omega)} \big). \end{align*} $ |
Similarly, we have, for $ t > 0 $,
| $ \begin{equation*} \|\nabla v_\varepsilon(t, \cdot) \|_{L^2(\Omega \setminus B_2)} \le Ce (\varepsilon, d) \big(\|f\|_{L^1\big( (0, + \infty); L^2(\Omega) \big)} +\|u_0\|_{L^2(\Omega)} \big). \end{equation*} $ |
The conclusion follows.
The second author is funded by the Vietnam National Foundation for Science and Technology Development (NAFOSTED) under grant number 101.02-2015.21.
The authors declare no conflict of interest in this paper.
| [1] |
The Mouse Genome Sequencing Consortium (2002) Initial sequencing and comparative analysis of the mouse genome. Nature 420: 520-562. doi: 10.1038/nature01262

|
| [2] | Woolfe A, Goodson M, Goode DK, et al. (2005) Highly conserved non-coding sequences are associated with vertebrate development. PLoS Biol 3: e7. |
| [3] |
Venkatesh B, Yap WH (2005) Comparative genomics using fugu: a tool for the identification of conserved vertebrate cis-regulatory elements. Bioessays 27: 100-107. doi: 10.1002/bies.20134
|
| [4] | Ahituv N, Prabhakar S, Poulin F, et al. (2005) Mapping cis-regulatory domains in the human genome using multi-species conservation of synteny. Hum Mol Genet 14: 3057-3063. |
| [5] | Shin JT, Priest JR, Ovcharenko I, et al. (2005) Human-zebrafish non-coding conserved elements act in vivo to regulate transcription. Nucleic Acids Res 33: 5437-5445. |
| [6] | Pennacchio LA, Ahituv N, Moses AM, et al. (2006) In vivo enhancer analysis of human conserved non-coding sequences. Nature 444: 499-502. |
| [7] | Allende ML, Manzanares M, Tena JJ, et al. (2006) Cracking the genome's second code: enhancer detection by combined phylogenetic foot-printing and transgenic fish and frog embryos. Methods 39: 212-219. |
| [8] | Berg JM, Tymoczko JL, Stryer L, et al. (2012) Biochemistry, 7th ed. WH Freeman & Company, New York. |
| [9] | Lodish H, Berk A, Kaiser C, et al. (2004) Molecular Cell Biology, 5th edition, WH Freeman and Company. |
| [10] | Cherstvy GA, Teif VB (2013) Structure-driven homology pairing of chromatin fibers: the role of electrostatics and protein-induced bridging. J Biol Phys 39: 363-385. |
| [11] |
Beshnova DA, Cherstvy AG, Vainshtein Y, et al. (2014) Regulation of the nucleosome repeat length in vivo by the DNA sequence, protein concentrations and long-range interactions. PLoS Comput Biol 10: e1003698. doi: 10.1371/journal.pcbi.1003698
|
| [12] | Sexton T, Cavalli G (2015) The Role of Chromosome Domains in Shaping the Functional Genome Cell 160: 1049-1059. |
| [13] | Rao SS, Huntley MH, Durand NC, et al. (2014) A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping. Cell 159: 1665-1680. |
| [14] | Jaenisch R, Bird A (2003) Epigenetic regulation of gene expression: how the genome integrates intrinsic and environmental signals. Nat Genetics 33: 245-254. |
| [15] | Kooistra SM, Helin K (2012) Molecular mechanisms and potential functions of histone demethylases. Nat Rev Mol Cell Biol 13: 297-311. |
| [16] | Kouzarides T (2007) Chromatin Modifications and Their Function. Cell 128: 693-705. |
| [17] | Barski A, Cuddapah S, Cui K, et al. (2007) High-resolution profiling of histone methylations in the human genome. Cell 129: 823-837. |
| [18] | Wu L, Wary KK, Revskoy S, et al. (2015) Histone Demethylases KDM4A and KDM4C Regulate Differentiation of Embryonic Stem Cells to Endothelial Cells. Stem Cell Reports Jun 24. pii: S2213-6711(15)00159-9. doi: 10.1016/j.stemcr.2015.05.016 [Epub ahead of print]. |
| [19] |
Kurdistani SK, Tavazoie S, Grunstein M (2004) Mapping global histone acetylation patterns to gene expression. Cell 117: 721-733. doi: 10.1016/j.cell.2004.05.023
|
| [20] |
Ganapathi M, Singh GP, Sandhu KS, et al. (2007) A whole genome analysis of 5' regulatory regions of human genes for putative cis-acting modulators of nucleosome positioning. Gene 391: 242-251. doi: 10.1016/j.gene.2007.01.008
|
| [21] |
Hebbar PB, Archer TK (2007) Chromatin-dependent cooperativity between site-specific transcription factors in vivo. J Biol Chem 282: 8284-8291. doi: 10.1074/jbc.M610554200
|
| [22] |
Gartenberg MR, Ampe C, Steitz TA, et al. (1990) Molecular characterization of the GCN4-DNA complex. Proc Natl Acad Sci U S A 87: 6034-6038. doi: 10.1073/pnas.87.16.6034
|
| [23] |
Dalma-Weiszhausz DD, Gartenberg MR, Crothers DM (1991) Sequence-dependent contribution of distal binding domains to CAP protein-DNA binding affinity. Nucleic Acids Res 19: 611-616. doi: 10.1093/nar/19.3.611
|
| [24] | Ulanovsky L, Bodner M, Trifonov EN, et al. (1986) Curved DNA: design, synthesis, and circularization. Proc Natl Acad Sci U S A 83: 862-866. |
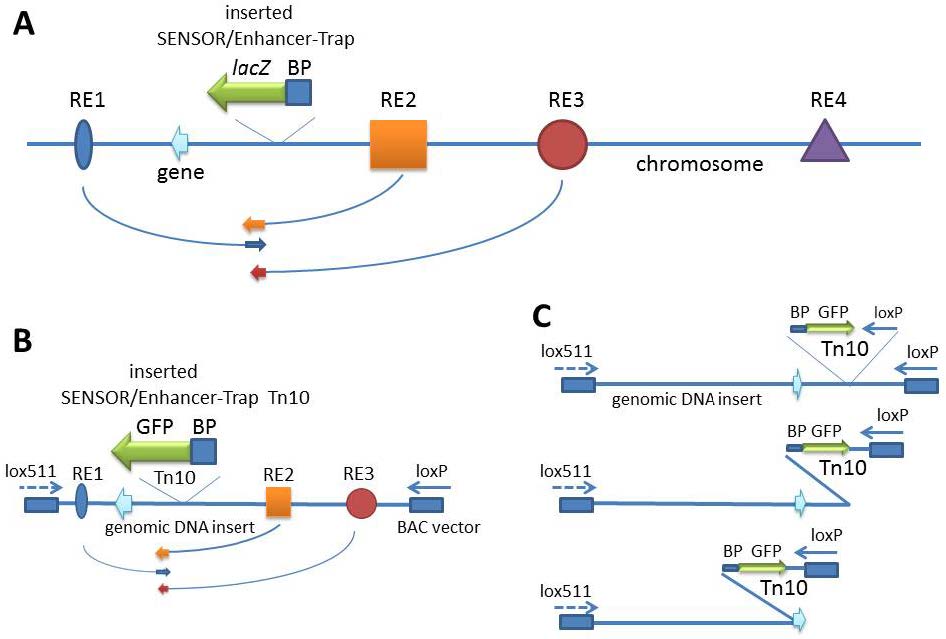
| [25] | Shakes LA, Du H, Wolf HM, et al. (2012) Using BAC Transgenesis in Zebrafish to Identify Regulatory sequences of the Amyloid Precursor Protein gene in Humans. BMC Genomics 13: 451. |
| [26] | Kawakami K, Takeda H, Kawakami N, et al. (2004) A transposon-mediated gene trap approach identifies developmentally regulated genes in zebrafish. Developmental Cell 7: 133-144. |
| [27] | Ellingsen S, Laplante MA, Konig M, et al. (2005) Large-scale enhancer detection in the zebrafish genome. Development 132: 3799-3811. |
| [28] |
Kokubu C, Horie K, Abe K, et al. (2009) A transposon-based chromosomal engineering method to survey a large cis-regulatory landscape in mice. Nat Genet 41: 946-952. doi: 10.1038/ng.397
|
| [29] | Ruf S, Symmons O, Uslu VV, et al. (2011) Large-scale analysis of the regulatory architecture of the mouse genome with a transposon-associated sensor. Nat Genet 43: 379-386. |
| [30] |
Symmons O, Spitz F (2013) From remote enhancers to gene regulation: charting the genome's regulatory landscapes. Philos Trans R Soc Lond B Biol Sci 368: 20120358. doi: 10.1098/rstb.2012.0358
|
| [31] | Bessa J, Tena JJ, de la Calle-Mustienes E, et al (2009) Zebrafish enhancer detection (ZED) vector: a new tool to facilitate transgenesis and the functional analysis of cis-regulatory regions in zebrafish. Dev Dyn 238: 2409-2417. |
| [32] | Chatterjee S, Lufkin T (2012) Regulatory genomics: Insights from the zebrafish. Curr Top Genet 5: 1-10. |
| [33] | Simon MI (1997) Dysfunctional genomics: BACs to the rescue. Nat Biotechnol 15: 839. |
| [34] |
Gilmore RC, Baker Jr J, Dempsey S, et al. (2001) Using PAC nested-deletions to order contigs and microsatellite markers at the high repetitive sequence containing Npr3 gene locus. Gene 275: 65-72. doi: 10.1016/S0378-1119(01)00654-0
|
| [35] | Shizuya H, Birren B, Kim UJ, et al. (1992) Cloning and stable maintenance of 300-kilobase-pair fragments of human DNA in Escherichia coli using an F-factor-based vector. Proc Natl Acad Sci U S A 89: 8794-8797. |
| [36] | Osoegawa K, Woon PY, Zhao B, et al. (1998) An improved approach for construction of bacterial artificial chromosome libraries. Genomics 52:1-8. |
| [37] | Frengen E, Weichenhan D, Zhao B, et al. (1999) A modular, positive selection bacterial artificial chromosome vector with multiple cloning sites. Genomics 58: 250-253. |
| [38] | Yang XW, Model P, Heintz N (1997) Homologous recombination based modification in Escherichia coli and germline transmission in transgenic mice of a bacterial artificial chromosome. Nat Biotechnol 9: 859-865 |
| [39] |
Gong S, Yang XW, Li C, et al. (2002) Highly efficient modification of bacterial artificial chromosomes (BACs) using novel shuttle vectors containing the R6Kgamma origin of replication. Genome Res 12: 1992-1998. doi: 10.1101/gr.476202
|
| [40] | Yang Z, Jiang H, Chachainasakul T, et al. (2006) Modified bacterial artificial chromosomes for zebrafish transgenesis. Methods 39:183-188. |
| [41] | Shakes LA, Wolf HM, Norford DC, et al. (2014) Harnessing Mobile Genetic Elements to Explore Gene Regulation. Mobile Genetic Elements 4: e29759. |
| [42] |
Zhang Y, Buchholz F, Muyrers JP, et al. (1998) A new logic for DNA engineering using recombination in Escherichia coli. Nat Genet 20: 123-128. doi: 10.1038/2417
|
| [43] |
Muyrers JP, Zhang Y, Testa G, et al. (1999) Rapid modification of Bacterial Artificial Chromosomes by ET recombination. Nucleic Acids Res 27: 1555-1557. doi: 10.1093/nar/27.6.1555
|
| [44] |
Suster ML, Abe G, Schouw A, et al. (2011) Transposon-mediated BAC transgenesis in zebrafish. Nat Protoc 6: 1998-2021 doi: 10.1038/nprot.2011.416
|
| [45] |
Warming S, Costantino N, Court DL, et al. (2005) Simple and highly efficient BAC recombineering using galK selection. Nucleic Acids Res 33: e36. doi: 10.1093/nar/gni035
|
| [46] | Fisher S, Grice EA, Vinton RM, et al. (2006) Conservation of RET regulatory function from human to zebrafish without sequence similarity. Science 14: 276-279. |
| [47] | Blow MJ, McCulley DJ, Li Z, et al. (2010) ChIP-Seq identification of weakly conserved heart enhancers. Nat Genet 42: 806-810. |
| [48] |
Kague E, Bessling SL, Lee J, et al. (2010) Functionally conserved cis-regulatory elements of COL18A1 identified through zebrafish transgenesis. Dev Biol 337: 496-505. doi: 10.1016/j.ydbio.2009.10.028
|
| [49] | Taher L, McGaughey DM, Maragh S, et al. (2011) Genome-wide identification of conserved regulatory function in diverged sequences. Genome Res 7: 1139-1149. |
| [50] |
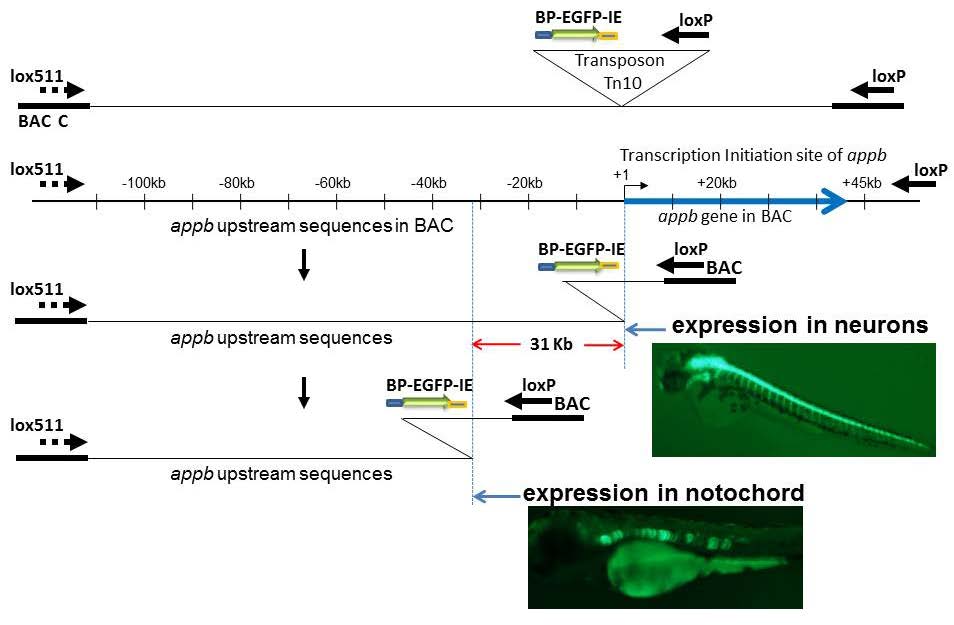
Shakes LA, Malcolm TL, Allen KL, et al. (2008) Context dependent function of APPb Enhancer identified using Enhancer Trap-containing BACs as Transgenes in Zebrafish. Nucleic Acids Res 36: 6237-6248. doi: 10.1093/nar/gkn628
|
| [51] | Chatterjee PK (2015) Directing Enhancer-traps and iTol2 End Sequences to Deleted BAC ends with loxP- and lox511-Tn10 transposons. “Bacterial Artificial Chromosomes” Methods in Molecular Biology series. Editor: K. Narayanan, Series editor: J. Walker. (2015) 2nd ed., Humana Press/ Springer, 99-122. |
| [52] |
Shakes LA, Garland DM, Srivastava DK, et al. (2005) Minimal Cross-recombination between wild type and loxP511 sites in vivo facilitates Truncating Both Ends of Large DNA Inserts in pBACe3.6 and Related Vectors. Nucleic Acids Res 33: e118. doi: 10.1093/nar/gni119
|
| [53] |
Shakes LA, Abe G, Eltayeb MA, et al. (2011) Generating libraries of iTol2-end insertions at BAC ends using loxP and lox511-Tn10 transposons. BMC Genomics 12: 351. doi: 10.1186/1471-2164-12-351
|
| [54] | Chatterjee PK, Shakes LA, Wolf HM, et al. (2013) Identifying Distal cis-acting Gene-Regulatory Sequences by Expressing BACs Functionalized with loxP-Tn10 Transposons in Zebrafish. Royal Soc Chem Adv 3: 8604-8617. |
| [55] | Mosimann C, Puller AC, Lawson KL, et al. (2013) Site-directed zebrafish transgenesis into single landing sites with the PhiC31 integrase system. Dev Dyn 242: 949-963. |
| [56] | Kirchmaier S, Höckendorf B, Möller EK, et al. (2013) Efficient site-specific transgenesis and enhancer activity tests in medaka using PhiC31 integrase. Development 140: 4287-4295. |
| [57] | Araki K, Araki M, Yamamura K (1997) Targeted integration of DNA using mutant lox sites in embryonic stem cells. Nucleic Acids Res 25: 868-872. |
| [58] | Liu W, Wang Y, Qin Y, et al. (2007) Site-Directed Gene Integration in Transgenic Zebrafish Mediated by Cre Recombinase Using a Combination of Mutant Lox Sites. Marine Biotechnol 9: 420-428. |
| [59] |
Musa A, Lehrach H, Russo VA (2001) Distinct expression patterns of two zebrafish homologues of the human APP gene during embryonic development. Dev Genes Evol 211: 563-567. doi: 10.1007/s00427-001-0189-9
|
| [60] | Salbaum JM, Weidemann A, Lemaire HG, et al. (1988) The promoter of Alzheimer's disease amyloid A4 precursor gene. EMBO J 7: 2807-2813. |
| [61] |
Yoshikai SI, Sasaki H, Dohura K, et al. (1990) Genomic organization of the human amyloid beta-protein precursor gene. Gene 87: 257-263. doi: 10.1016/0378-1119(90)90310-N
|
| [62] |
Lahiri DK, Robakis NK (1991) The promoter activity of the gene encoding Alzheimer beta-amyloid precursor protein (APP) is regulated by two blocks of upstream sequences. Brain Res Mol Brain Res 9: 253-257. doi: 10.1016/0169-328X(91)90009-M
|
| [63] |
Song W, Lahiri DK (1998) Functional identification of the promoter of the gene encoding the Rhesus monkey beta-amyloid precursor protein. Gene 217:165-176. doi: 10.1016/S0378-1119(98)00340-0
|
| [64] | Lahiri DK, Song W, Ge YW (2000) Analysis of the 5'-flanking region of the beta-amyloid precursor protein gene that contributes to increased promoter activity in differentiated neuronal cells, Brain Res Mol Brain Res 77: 185-198. |
| [65] | Lahiri DK, Ge YW, Maloney B (2005) Characterization of the APP proximal promoter and 5'-untranslated regions: Identification of cell-type specific domains and implications in APP gene expression and Alzheimer's disease. FASEB J 19: 653-665. |
| [66] | Rogers JT, Randall JD, Cahill CM, et al. (2002) An iron-responsive element type II in the 5' untranslated region of the Alzheimer's amyloid precursor protein transcript. J Biol Chem 277: 518-528. |
| [67] |
Shaw KT, Utsuki T, Rogers J, et al. (2001) Phenserine regulates translation of beta-amyloid precursor protein mRNA by a putative interleukin-1 responsive element, a target for drug development. Proc Natl Acad Sci U S A 98: 7605-7610. doi: 10.1073/pnas.131152998
|
| [68] |
Theuns J, Brouwers N, Engelborghs S, et al. (2006) Promoter mutations that increase amyloid precursor-protein expression are associated with Alzheimer disease. Am J Hum Genet 78: 936-946. doi: 10.1086/504044
|
| [69] |
Chakraborty T, Perlot T, Subrahmanyam R, et al. (2009) A 220-nucleotide deletion of the intronic enhancer reveals an epigenetic hierarchy in immunoglobulin heavy chain locus activation. J Exp Med 206: 1019-1027. doi: 10.1084/jem.20081621
|
| [70] |
Wolf HM, Nyabera KO, De La Torre KK, et al. (2014) Long Range Gene-Regulatory Sequences Identified by Transgenic Expression of Bacterially-Engineered Enhancer-trap BACs in Zebrafish. Mol Biol Genetic Eng 2: 2. doi: 10.7243/2053-5767-2-2
|
| [71] |
Popović M, Caballero-Bleda M, Puelles L, et al. (1998) Importance of immunological and inflammatory processes in the pathogenesis and therapy of Alzheimer's disease. Int J Neurosci 95: 203-236. doi: 10.3109/00207459809003341
|
| [72] |
Kamizono S, Duncan GS, Seidel MG, et al. (2009) Nfil3/E4bp4 is required for the development and maturation of NK cells in vivo. J Exp Med 206: 2977-2986. doi: 10.1084/jem.20092176
|
| [73] |
Kobayashi T, Matsuoka K, Sheikh SZ, et al. (2011) NFIL3 is a regulator of IL-12 p40 in macrophages and mucosal immunity. J Immunol 186: 4649-4655. doi: 10.4049/jimmunol.1003888
|
| [74] |
The ENCODE Project Consortium (2007) Identification and analysis of functional elements in 1% of the human genome by the ENCODE pilot project. Nature 447: 799-816. doi: 10.1038/nature05874
|
| [75] |
The International HapMap 3 Consortium (2010) Integrating common and rare genetic variation in diverse human populations. Nature 467: 52-58. doi: 10.1038/nature09298
|
| [76] |
The 1000 Genomes Project Consortium (2012) An integrated map of genetic variation from 1,092 human genomes. Nature 491: 56-65. doi: 10.1038/nature11632
|
| 1. | Vladimir B. Teif, Andrey G. Cherstvy, Chromatin and epigenetics: current biophysical views, 2016, 3, 2377-9098, 88, 10.3934/biophy.2016.1.88 |
Figures(4)

Delores J. Grant, Leighcraft A. Shakes, Hope M. Wolf, Derek C. Norford, Pradeep K. Chatterjee. Exploring function of conserved non-coding DNA in its chromosomal context[J]. AIMS Biophysics, 2015, 2(4): 773-793. doi: 10.3934/biophy.2015.4.773







 DownLoad:
DownLoad: