Citation: Chao Hu, Qing-Hua Qin. Bone remodeling and biological effects of mechanical stimulus[J]. AIMS Bioengineering, 2020, 7(1): 12-28. doi: 10.3934/bioeng.2020002

| [1] |
Hu C, Ashok D, Nisbet DR, et al. (2019) Bioinspired surface modification of orthopedic implants for bone tissue engineering. Biomaterials 119366. doi: 10.1016/j.biomaterials.2019.119366

|
| [2] |
Karsenty G, Olson EN (2016) Bone and muscle endocrine functions: unexpected paradigms of inter-organ communication. Cell 164: 1248-1256. doi: 10.1016/j.cell.2016.02.043
|
| [3] |
Rossi M, Battafarano G, Pepe J, et al. (2019) The endocrine function of osteocalcin regulated by bone resorption: A lesson from reduced and increased bone mass diseases. Int J Mol Sci 20: 4502. doi: 10.3390/ijms20184502
|
| [4] |
Loebel C, Burdick JA (2018) Engineering stem and stromal cell therapies for musculoskeletal tissue repair. Cell Stem Cell 22: 325-339. doi: 10.1016/j.stem.2018.01.014
|
| [5] |
Dimitriou R, Jones E, McGonagle D, et al. (2011) Bone regeneration: current concepts and future directions. BMC Med 9: 66. doi: 10.1186/1741-7015-9-66
|
| [6] | Nordin M, Frankel VH (2001) Basic Biomechanics of the Musculoskeletal System, 3 Eds USA: Lippincott Williams & Wilkins. |
| [7] |
Kobayashi S, Takahashi HE, Ito A, et al. (2003) Trabecular minimodeling in human iliac bone. Bone 32: 163-169. doi: 10.1016/S8756-3282(02)00947-X
|
| [8] |
Bartl R, Bartl C (2019) Control and regulation of bone remodelling. The Osteoporosis Manual Cham: Springer, 31-39. doi: 10.1007/978-3-030-00731-7_4
|
| [9] |
Kenkre JS, Bassett JHD (2018) The bone remodelling cycle. Ann Clin Biochem 55: 308-327. doi: 10.1177/0004563218759371
|
| [10] |
Prendergast PJ, Huiskes R (1995) The biomechanics of Wolff's law: recent advances. Irish J Med Sci 164: 152-154. doi: 10.1007/BF02973285
|
| [11] |
Wegst UGK, Bai H, Saiz E, et al. (2015) Bioinspired structural materials. Nat Mater 14: 23-36. doi: 10.1038/nmat4089
|
| [12] |
Reznikov N, Shahar R, Weiner S (2014) Bone hierarchical structure in three dimensions. Acta Biomater 10: 3815-3826. doi: 10.1016/j.actbio.2014.05.024
|
| [13] |
Weiner S, Wagner HD (1998) The material bone: structure-mechanical function relations. Annu Rev Mater Sci 28: 271-298. doi: 10.1146/annurev.matsci.28.1.271
|
| [14] |
Recker RR, Kimmel DB, Dempster D, et al. (2011) Issues in modern bone histomorphometry. Bone 49: 955-964. doi: 10.1016/j.bone.2011.07.017
|
| [15] |
Eriksen EF, Vesterby A, Kassem M, et al. (1993) Bone remodeling and bone structure. Physiology and Pharmacology of Bone Heidelberg: Springer, 67-109. doi: 10.1007/978-3-642-77991-6_2
|
| [16] |
Augat P, Schorlemmer S (2006) The role of cortical bone and its microstructure in bone strength. Age Ageing 35: ii27-ii31. doi: 10.1093/ageing/afl081
|
| [17] |
Kozielski M, Buchwald T, Szybowicz M, et al. (2011) Determination of composition and structure of spongy bone tissue in human head of femur by Raman spectral mapping. J Mater Sci: Mater Med 22: 1653-1661. doi: 10.1007/s10856-011-4353-0
|
| [18] |
Cross LM, Thakur A, Jalili NA, et al. (2016) Nanoengineered biomaterials for repair and regeneration of orthopedic tissue interfaces. Acta Biomater 42: 2-17. doi: 10.1016/j.actbio.2016.06.023
|
| [19] |
Zebaze R, Seeman E (2015) Cortical bone: a challenging geography. J Bone Miner Res 30: 24-29. doi: 10.1002/jbmr.2419
|
| [20] |
Liu Y, Luo D, Wang T (2016) Hierarchical structures of bone and bioinspired bone tissue engineering. Small 12: 4611-4632. doi: 10.1002/smll.201600626
|
| [21] |
Brodsky B, Persikov AV (2005) Molecular structure of the collagen triple helix. Adv Protein Chem 70: 301-339. doi: 10.1016/S0065-3233(05)70009-7
|
| [22] |
Cui FZ, Li Y, Ge J (2007) Self-assembly of mineralized collagen composites. Mater Sci Eng R Rep 57: 1-27. doi: 10.1016/j.mser.2007.04.001
|
| [23] |
Wang Y, Azaïs T, Robin M, et al. (2012) The predominant role of collagen in the nucleation, growth, structure and orientation of bone apatite. Nat Mater 11: 724-733. doi: 10.1038/nmat3362
|
| [24] | Bentmann A, Kawelke N, Moss D, et al. (2010) Circulating fibronectin affects bone matrix, whereas osteoblast fibronectin modulates osteoblast function. J Bone Miner Res 25: 706-715. |
| [25] |
Szweras M, Liu D, Partridge EA, et al. (2002) α2-HS glycoprotein/fetuin, a transforming growth factor-β/bone morphogenetic protein antagonist, regulates postnatal bone growth and remodeling. J Biol Chem 277: 19991-19997. doi: 10.1074/jbc.M112234200
|
| [26] |
Boskey AL, Robey PG (2013) The regulatory role of matrix proteins in mineralization of bone. Osteoporosis, 4 Eds Academic Press, 235-255. doi: 10.1016/B978-0-12-415853-5.00011-X
|
| [27] |
Boskey AL (2013) Bone composition: relationship to bone fragility and antiosteoporotic drug effects. Bonekey Rep 2: 447. doi: 10.1038/bonekey.2013.181
|
| [28] |
Stock SR (2015) The mineral–collagen interface in bone. Calcified Tissue Int 97: 262-280. doi: 10.1007/s00223-015-9984-6
|
| [29] |
Nikel O, Laurencin D, McCallum SA, et al. (2013) NMR investigation of the role of osteocalcin and osteopontin at the organic–inorganic interface in bone. Langmuir 29: 13873-13882. doi: 10.1021/la403203w
|
| [30] |
He G, Dahl T, Veis A, et al. (2003) Nucleation of apatite crystals in vitro by self-assembled dentin matrix protein 1. Nat Mater 2: 552-558. doi: 10.1038/nmat945
|
| [31] |
Clarke B (2008) Normal bone anatomy and physiology. Clin J Am Soc Nephro 3: S131-S139. doi: 10.2215/CJN.04151206
|
| [32] |
Olszta MJ, Cheng X, Jee SS, et al. (2007) Bone structure and formation: A new perspective. Mater Sci Eng R Rep 58: 77-116. doi: 10.1016/j.mser.2007.05.001
|
| [33] |
Nair AK, Gautieri A, Chang SW, et al. (2013) Molecular mechanics of mineralized collagen fibrils in bone. Nature Commun 4: 1724. doi: 10.1038/ncomms2720
|
| [34] |
Landis WJ (1995) The strength of a calcified tissue depends in part on the molecular structure and organization of its constituent mineral crystals in their organic matrix. Bone 16: 533-544. doi: 10.1016/8756-3282(95)00076-P
|
| [35] |
Hunter GK, Hauschka PV, POOLE RA, et al. (1996) Nucleation and inhibition of hydroxyapatite formation by mineralized tissue proteins. Biochem J 317: 59-64. doi: 10.1042/bj3170059
|
| [36] |
Oikeh I, Sakkas P, Blake D P, et al. (2019) Interactions between dietary calcium and phosphorus level, and vitamin D source on bone mineralization, performance, and intestinal morphology of coccidia-infected broilers. Poult Sci 11: 5679-5690. doi: 10.3382/ps/pez350
|
| [37] |
Boyce BF, Rosenberg E, de Papp AE, et al. (2012) The osteoclast, bone remodelling and treatment of metabolic bone disease. Eur J Clin Invest 42: 1332-1341. doi: 10.1111/j.1365-2362.2012.02717.x
|
| [38] |
Teitelbaum SL (2000) Bone resorption by osteoclasts. Science 289: 1504-1508. doi: 10.1126/science.289.5484.1504
|
| [39] |
Yoshida H, Hayashi SI, Kunisada T, et al. (1990) The murine mutation osteopetrosis is in the coding region of the macrophage colony stimulating factor gene. Nature 345: 442-444. doi: 10.1038/345442a0
|
| [40] |
Roodman GD (2006) Regulation of osteoclast differentiation. Ann NY Acad Sci 1068: 100-109. doi: 10.1196/annals.1346.013
|
| [41] |
Martin TJ (2013) Historically significant events in the discovery of RANK/RANKL/OPG. World J Orthop 4: 186-197. doi: 10.5312/wjo.v4.i4.186
|
| [42] |
Coetzee M, Haag M, Kruger MC (2007) Effects of arachidonic acid, docosahexaenoic acid, prostaglandin E2 and parathyroid hormone on osteoprotegerin and RANKL secretion by MC3T3-E1 osteoblast-like cells. J Nutr Biochem 18: 54-63. doi: 10.1016/j.jnutbio.2006.03.002
|
| [43] |
Steeve KT, Marc P, Sandrine T, et al. (2004) IL-6, RANKL, TNF-alpha/IL-1: interrelations in bone resorption pathophysiology. Cytokine Growth F R 15: 49-60. doi: 10.1016/j.cytogfr.2003.10.005
|
| [44] |
Mellis DJ, Itzstein C, Helfrich M, et al. (2011) The skeleton: a multi-functional complex organ. The role of key signalling pathways in osteoclast differentiation and in bone resorption. J Endocrinol 211: 131-143. doi: 10.1530/JOE-11-0212
|
| [45] | Silva I, Branco J (2011) Rank/Rankl/opg: literature review. Acta Reumatol Port 36: 209-218. |
| [46] |
Martin TJ, Sims NA (2015) RANKL/OPG; Critical role in bone physiology. Rev Endocr Metab Dis 16: 131-139. doi: 10.1007/s11154-014-9308-6
|
| [47] |
Wang Y, Qin QH (2012) A theoretical study of bone remodelling under PEMF at cellular level. Comput Method Biomec 15: 885-897. doi: 10.1080/10255842.2011.565752
|
| [48] |
Weitzmann MN, Pacifici R (2007) T cells: unexpected players in the bone loss induced by estrogen deficiency and in basal bone homeostasis. Ann NY Acad Sci 1116: 360-375. doi: 10.1196/annals.1402.068
|
| [49] |
Duong LT, Lakkakorpi P, Nakamura I, et al. (2000) Integrins and signaling in osteoclast function. Matrix Biol 19: 97-105. doi: 10.1016/S0945-053X(00)00051-2
|
| [50] |
Stenbeck G (2002) Formation and function of the ruffled border in osteoclasts. Semin Cell Dev Biol 13: 285-292. doi: 10.1016/S1084952102000587
|
| [51] |
Jurdic P, Saltel F, Chabadel A, et al. (2006) Podosome and sealing zone: specificity of the osteoclast model. Eur J Cell Biol 85: 195-202. doi: 10.1016/j.ejcb.2005.09.008
|
| [52] |
Väänänen HK, Laitala-Leinonen T (2008) Osteoclast lineage and function. Arch Biochem Biophys 473: 132-138. doi: 10.1016/j.abb.2008.03.037
|
| [53] | Vaananen HK, Zhao H, Mulari M, et al. (2000) The cell biology of osteoclast function. J cell Sci 113: 377-381. |
| [54] |
Sabolová V, Brinek A, Sládek V (2018) The effect of hydrochloric acid on microstructure of porcine (Sus scrofa domesticus) cortical bone tissue. Forensic Sci Int 291: 260-271. doi: 10.1016/j.forsciint.2018.08.030
|
| [55] |
Delaissé JM, Engsig MT, Everts V, et al. (2000) Proteinases in bone resorption: obvious and less obvious roles. Clin Chim Acta 291: 223-234. doi: 10.1016/S0009-8981(99)00230-2
|
| [56] |
Logar DB, Komadina R, Preželj J, et al. (2007) Expression of bone resorption genes in osteoarthritis and in osteoporosis. J Bone Miner Metab 25: 219-225. doi: 10.1007/s00774-007-0753-0
|
| [57] |
Lorget F, Kamel S, Mentaverri R, et al. (2000) High extracellular calcium concentrations directly stimulate osteoclast apoptosis. Biochem Bioph Res Co 268: 899-903. doi: 10.1006/bbrc.2000.2229
|
| [58] |
Nesbitt SA, Horton MA (1997) Trafficking of matrix collagens through bone-resorbing osteoclasts. Science 276: 266-269. doi: 10.1126/science.276.5310.266
|
| [59] |
Xing L, Boyce BF (2005) Regulation of apoptosis in osteoclasts and osteoblastic cells. Biochem Bioph Res Co 328: 709-720. doi: 10.1016/j.bbrc.2004.11.072
|
| [60] |
Hughes DE, Wright KR, Uy HL, et al. (1995) Bisphosphonates promote apoptosis in murine osteoclasts in vitro and in vivo. J Bone Miner Res 10: 1478-1487. doi: 10.1002/jbmr.5650101008
|
| [61] |
Choi Y, Arron JR, Townsend MJ (2009) Promising bone-related therapeutic targets for rheumatoid arthritis. Nat Rev Rheumatol 5: 543-548. doi: 10.1038/nrrheum.2009.175
|
| [62] |
Harvey NC, McCloskey E, Kanis JA, et al. (2017) Bisphosphonates in osteoporosis: NICE and easy? Lancet 390: 2243-2244. doi: 10.1016/S0140-6736(17)32850-7
|
| [63] |
Ducy P, Schinke T, Karsenty G (2000) The osteoblast: a sophisticated fibroblast under central surveillance. Science 289: 1501-1504. doi: 10.1126/science.289.5484.1501
|
| [64] |
Katagiri T, Takahashi N (2002) Regulatory mechanisms of osteoblast and osteoclast differentiation. Oral dis 8: 147-159. doi: 10.1034/j.1601-0825.2002.01829.x
|
| [65] |
Kretzschmar M, Liu F, Hata A, et al. (1997) The TGF-beta family mediator Smad1 is phosphorylated directly and activated functionally by the BMP receptor kinase. Gene Dev 11: 984-995. doi: 10.1101/gad.11.8.984
|
| [66] |
Bennett CN, Longo KA, Wright WS, et al. (2005) Regulation of osteoblastogenesis and bone mass by Wnt10b. P Natl A Sci 102: 3324-3329. doi: 10.1073/pnas.0408742102
|
| [67] | Wang Y, Qin QH, Kalyanasundaram S (2009) A theoretical model for simulating effect of parathyroid hormone on bone metabolism at cellular level. Mol Cell Biomech 6: 101-112. |
| [68] |
Elefteriou F, Ahn JD, Takeda S, et al. (2005) Leptin regulation of bone resorption by the sympathetic nervous system and CART. Nature 434: 514-520. doi: 10.1038/nature03398
|
| [69] |
Proff P, Römer P (2009) The molecular mechanism behind bone remodelling: a review. Clin Oral Invest 13: 355-362. doi: 10.1007/s00784-009-0268-2
|
| [70] |
Katsimbri P (2017) The biology of normal bone remodelling. Eur J Cancer Care 26: e12740. doi: 10.1111/ecc.12740
|
| [71] |
Fratzl P, Weinkamer R (2007) Nature's hierarchical materials. Prog Mater Sci 52: 1263-1334. doi: 10.1016/j.pmatsci.2007.06.001
|
| [72] |
Athanasiou KA, Zhu CF, Lanctot DR, et al. (2000) Fundamentals of biomechanics in tissue engineering of bone. Tissue Eng 6: 361-381. doi: 10.1089/107632700418083
|
| [73] |
Takahashi N, Udagawa N, Suda T (1999) A new member of tumor necrosis factor ligand family, ODF/OPGL/TRANCE/RANKL, regulates osteoclast differentiation and function. Biocheml Bioph Res Co 256: 449-455. doi: 10.1006/bbrc.1999.0252
|
| [74] |
Nakashima T, Hayashi M, Fukunaga T, et al. (2011) Evidence for osteocyte regulation of bone homeostasis through RANKL expression. Nat Med 17: 1231-1234. doi: 10.1038/nm.2452
|
| [75] |
Prideaux M, Findlay DM, Atkins GJ (2016) Osteocytes: the master cells in bone remodelling. Curr Opin Pharmacol 28: 24-30. doi: 10.1016/j.coph.2016.02.003
|
| [76] |
Dallas SL, Prideaux M, Bonewald LF (2013) The osteocyte: an endocrine cell… and more. Endocr Rev 34: 658-690. doi: 10.1210/er.2012-1026
|
| [77] |
Rochefort GY, Pallu S, Benhamou CL (2010) Osteocyte: the unrecognized side of bone tissue. Osteoporosis Int 21: 1457-1469. doi: 10.1007/s00198-010-1194-5
|
| [78] |
Rowe PSN (2012) Regulation of bone–renal mineral and energy metabolism: The PHEX, FGF23, DMP1, MEPE ASARM pathway. Crit Rev Eukaryot Gene Expr 22: 61-86. doi: 10.1615/CritRevEukarGeneExpr.v22.i1.50
|
| [79] |
Pajevic PD, Krause DS (2019) Osteocyte regulation of bone and blood. Bone 119: 13-18. doi: 10.1016/j.bone.2018.02.012
|
| [80] | Frost HM (1987) The mechanostat: a proposed pathogenic mechanism of osteoporoses and the bone mass effects of mechanical and nonmechanical agents. Bone Miner 2: 73-85. |
| [81] |
Tate MLK, Adamson JR, Tami AE, et al. (2004) The osteocyte. Int J Biochem Cell Biol 36: 1-8. doi: 10.1016/S1357-2725(03)00241-3
|
| [82] |
Bonewald LF, Johnson ML (2008) Osteocytes, mechanosensing and Wnt signaling. Bone 42: 606-615. doi: 10.1016/j.bone.2007.12.224
|
| [83] |
Manolagas SC, Parfitt AM (2010) What old means to bone. Trends Endocrinol Metab 21: 369-374. doi: 10.1016/j.tem.2010.01.010
|
| [84] |
Wang Y, Qin QH (2010) Parametric study of control mechanism of cortical bone remodeling under mechanical stimulus. Acta Mech Sinica 26: 37-44. doi: 10.1007/s10409-009-0313-z
|
| [85] |
Qu C, Qin QH, Kang Y (2006) A hypothetical mechanism of bone remodeling and modeling under electromagnetic loads. Biomaterials 27: 4050-4057. doi: 10.1016/j.biomaterials.2006.03.015
|
| [86] |
Parfitt AM (2002) Targeted and nontargeted bone remodeling: relationship to basic multicellular unit origination and progression. Bone 1: 5-7. doi: 10.1016/S8756-3282(01)00642-1
|
| [87] |
Hadjidakis DJ, Androulakis II (2006) Bone remodeling. Ann NYAcad Sci 1092: 385-396. doi: 10.1196/annals.1365.035
|
| [88] | Vaananen HK, Zhao H, Mulari M, et al. (2000) The cell biology of osteoclast function. J cell Sci 113: 377-381. |
| [89] |
Goldring SR (2015) The osteocyte: key player in regulating bone turnover. RMD Open 1: e000049. doi: 10.1136/rmdopen-2015-000049
|
| [90] |
Silver IA, Murrills RJ, Etherington DJ (1988) Microelectrode studies on the acid microenvironment beneath adherent macrophages and osteoclasts. Exp Cell Res 175: 266-276. doi: 10.1016/0014-4827(88)90191-7
|
| [91] |
Delaissé JM, Andersen TL, Engsig MT, et al. (2003) Matrix metalloproteinases (MMP) and cathepsin K contribute differently to osteoclastic activities. Microsc Res Techniq 61: 504-513. doi: 10.1002/jemt.10374
|
| [92] |
Delaisse JM (2014) The reversal phase of the bone-remodeling cycle: cellular prerequisites for coupling resorption and formation. Bonekey Rep 3: 561. doi: 10.1038/bonekey.2014.56
|
| [93] | Bonewald LF, Mundy GR (1990) Role of transforming growth factor-beta in bone remodeling. Clin Orthop Relat R 250: 261-276. |
| [94] |
Locklin RM, Oreffo ROC, Triffitt JT (1999) Effects of TGFβ and bFGF on the differentiation of human bone marrow stromal fibroblasts. Cell Biol Int 23: 185-194. doi: 10.1006/cbir.1998.0338
|
| [95] |
Lee B, Oh Y, Jo S, et al. (2019) A dual role of TGF-β in human osteoclast differentiation mediated by Smad1 versus Smad3 signaling. Immunol Lett 206: 33-40. doi: 10.1016/j.imlet.2018.12.003
|
| [96] |
Koseki T, Gao Y, Okahashi N, et al. (2002) Role of TGF-β family in osteoclastogenesis induced by RANKL. Cell Signal 14: 31-36. doi: 10.1016/S0898-6568(01)00221-2
|
| [97] |
Anderson HC (2003) Matrix vesicles and calcification. Curr Rheumatol Rep 5: 222-226. doi: 10.1007/s11926-003-0071-z
|
| [98] |
Bellido T, Plotkin LI, Bruzzaniti A (2019) Bone cells. Basic and Applied Bone Biology, 2 Eds Elsevier, 37-55. doi: 10.1016/B978-0-12-813259-3.00003-8
|
| [99] |
Weinstein RS, Jilka RL, Parfitt AM, et al. (1998) Inhibition of osteoblastogenesis and promotion of apoptosis of osteoblasts and osteocytes by glucocorticoids. Potential mechanisms of their deleterious effects on bone. J Clin Invest 102: 274-282. doi: 10.1172/JCI2799
|
| [100] |
Vezeridis PS, Semeins CM, Chen Q, et al. (2006) Osteocytes subjected to pulsating fluid flow regulate osteoblast proliferation and differentiation. Biochem Bioph Res Co 348: 1082-1088. doi: 10.1016/j.bbrc.2006.07.146
|
| [101] |
Lind M, Deleuran B, Thestrup-Pedersen K, et al. (1995) Chemotaxis of human osteoblasts: Effects of osteotropic growth factors. Apmis 103: 140-146. doi: 10.1111/j.1699-0463.1995.tb01089.x
|
| [102] |
Russo CR, Lauretani F, Seeman E, et al. (2006) Structural adaptations to bone loss in aging men and women. Bone 38: 112-118. doi: 10.1016/j.bone.2005.07.025
|
| [103] |
Ozcivici E, Luu YK, Adler B, et al. (2010) Mechanical signals as anabolic agents in bone. Nat Rev Rheumatol 6: 50-59. doi: 10.1038/nrrheum.2009.239
|
| [104] |
Rosa N, Simoes R, Magalhães FD, et al. (2015) From mechanical stimulus to bone formation: a review. Med Eng Phys 37: 719-728. doi: 10.1016/j.medengphy.2015.05.015
|
| [105] |
Noble BS, Peet N, Stevens HY, et al. (2003) Mechanical loading: biphasic osteocyte survival and targeting of osteoclasts for bone destruction in rat cortical bone. Am J Physiol-Cell Ph 284: C934-C943. doi: 10.1152/ajpcell.00234.2002
|
| [106] |
Robling AG, Castillo AB, Turner CH (2006) Biomechanical and molecular regulation of bone remodeling. Annu Rev Biomed Eng 8: 455-498. doi: 10.1146/annurev.bioeng.8.061505.095721
|
| [107] |
Qin QH, Mai YW (1999) A closed crack tip model for interface cracks inthermopiezoelectric materials. Int J Solids Struct 36: 2463-2479. doi: 10.1016/S0020-7683(98)00115-2
|
| [108] |
Yu SW, Qin QH (1996) Damage analysis of thermopiezoelectric properties: Part I—crack tip singularities. Theor Appl Fract Mec 25: 263-277. doi: 10.1016/S0167-8442(96)00026-2
|
| [109] |
Qin QH, Mai YW, Yu SW (1998) Effective moduli for thermopiezoelectric materials with microcracks. Int J Fracture 91: 359-371. doi: 10.1023/A:1007423508650
|
| [110] |
Jirousek J, Qin QH (1996) Application of hybrid-Trefftz element approach to transient heat conduction analysis. Comput Struct 58: 195-201. doi: 10.1016/0045-7949(95)00115-W
|
| [111] |
Qin QH (1995) Hybrid-Trefftz finite element method for Reissner plates on an elastic foundation. Comput Method Appl M 122: 379-392. doi: 10.1016/0045-7825(94)00730-B
|
| [112] |
Qin QH (1994) Hybrid Trefftz finite-element approach for plate bending on an elastic foundation. Appl Math Model 18: 334-339. doi: 10.1016/0307-904X(94)90357-3
|
| [113] |
Qin QH (2013) Mechanics of Cellular Bone Remodeling: Coupled Thermal, Electrical, and Mechanical Field Effects CRC Press. doi: 10.1201/b13728
|
| [114] | Wang H, Qin QH (2010) FE approach with Green's function as internal trial function for simulating bioheat transfer in the human eye. Arch Mech 62: 493-510. |
| [115] | Qin QH (2003) Fracture analysis of cracked thermopiezoelectric materials by BEM. Electronic J Boundary Elem 1: 283-301. |
| [116] |
Qin QH, Ye JQ (2004) Thermoelectroelastic solutions for internal bone remodeling under axial and transverse loads. Int J Solids Struct 41: 2447-2460. doi: 10.1016/j.ijsolstr.2003.12.026
|
| [117] |
Qin QH, Qu C, Ye J (2005) Thermoelectroelastic solutions for surface bone remodeling under axial and transverse loads. Biomaterials 26: 6798-6810. doi: 10.1016/j.biomaterials.2005.03.042
|
| [118] |
Ducher G, Jaffré C, Arlettaz A, et al. (2005) Effects of long-term tennis playing on the muscle-bone relationship in the dominant and nondominant forearms. Can J Appl Physiol 30: 3-17. doi: 10.1139/h05-101
|
| [119] |
Robling AG, Hinant FM, Burr DB, et al. (2002) Improved bone structure and strength after long-term mechanical loading is greatest if loading is separated into short bouts. J Bone Miner Res 17: 1545-1554. doi: 10.1359/jbmr.2002.17.8.1545
|
| [120] |
Rubin J, Rubin C, Jacobs CR (2006) Molecular pathways mediating mechanical signaling in bone. Gene 367: 1-16. doi: 10.1016/j.gene.2005.10.028
|
| [121] |
Tatsumi S, Ishii K, Amizuka N, et al. (2007) Targeted ablation of osteocytes induces osteoporosis with defective mechanotransduction. Cell Metab 5: 464-475. doi: 10.1016/j.cmet.2007.05.001
|
| [122] |
Robling AG, Turner CH (2009) Mechanical signaling for bone modeling and remodeling. Crit Rev Eukar Gene 19: 319-338. doi: 10.1615/CritRevEukarGeneExpr.v19.i4.50
|
| [123] |
Galli C, Passeri G, Macaluso GM (2010) Osteocytes and WNT: the mechanical control of bone formation. J Dent Res 89: 331-343. doi: 10.1177/0022034510363963
|
| [124] |
Robling AG, Duijvelaar KM, Geevers JV, et al. (2001) Modulation of appositional and longitudinal bone growth in the rat ulna by applied static and dynamic force. Bone 29: 105-113. doi: 10.1016/S8756-3282(01)00488-4
|
| [125] |
Burr DB, Milgrom C, Fyhrie D, et al. (1996) In vivo measurement of human tibial strains during vigorous activity. Bone 18: 405-410. doi: 10.1016/8756-3282(96)00028-2
|
| [126] |
Sun W, Chi S, Li Y, et al. (2019) The mechanosensitive Piezo1 channel is required for bone formation. Elife 8: e47454. doi: 10.7554/eLife.47454
|
| [127] |
Goda I, Ganghoffer JF, Czarnecki S, et al. (2019) Topology optimization of bone using cubic material design and evolutionary methods based on internal remodeling. Mech Res Commun 95: 52-60. doi: 10.1016/j.mechrescom.2018.12.003
|
| [128] |
Goda I, Ganghoffer JF (2018) Modeling of anisotropic remodeling of trabecular bone coupled to fracture. Arch Appl Mech 88: 2101-2121. doi: 10.1007/s00419-018-1438-y
|
| [129] |
Louna Z, Goda I, Ganghoffer JF, et al. (2017) Formulation of an effective growth response of trabecular bone based on micromechanical analyses at the trabecular level. Arch Appl Mech 87: 457-477. doi: 10.1007/s00419-016-1204-y
|
| [130] |
Goda I, Ganghoffer JF (2017) Construction of the effective plastic yield surfaces of vertebral trabecular bone under twisting and bending moments stresses using a 3D microstructural model. ZAMM Z Angew Math Mech 97: 254-272. doi: 10.1002/zamm.201600141
|
| [131] |
Qin QH, Wang YN (2012) A mathematical model of cortical bone remodeling at cellular level under mechanical stimulus. Acta Mech Sinica-Prc 28: 1678-1692. doi: 10.1007/s10409-012-0154-z
|
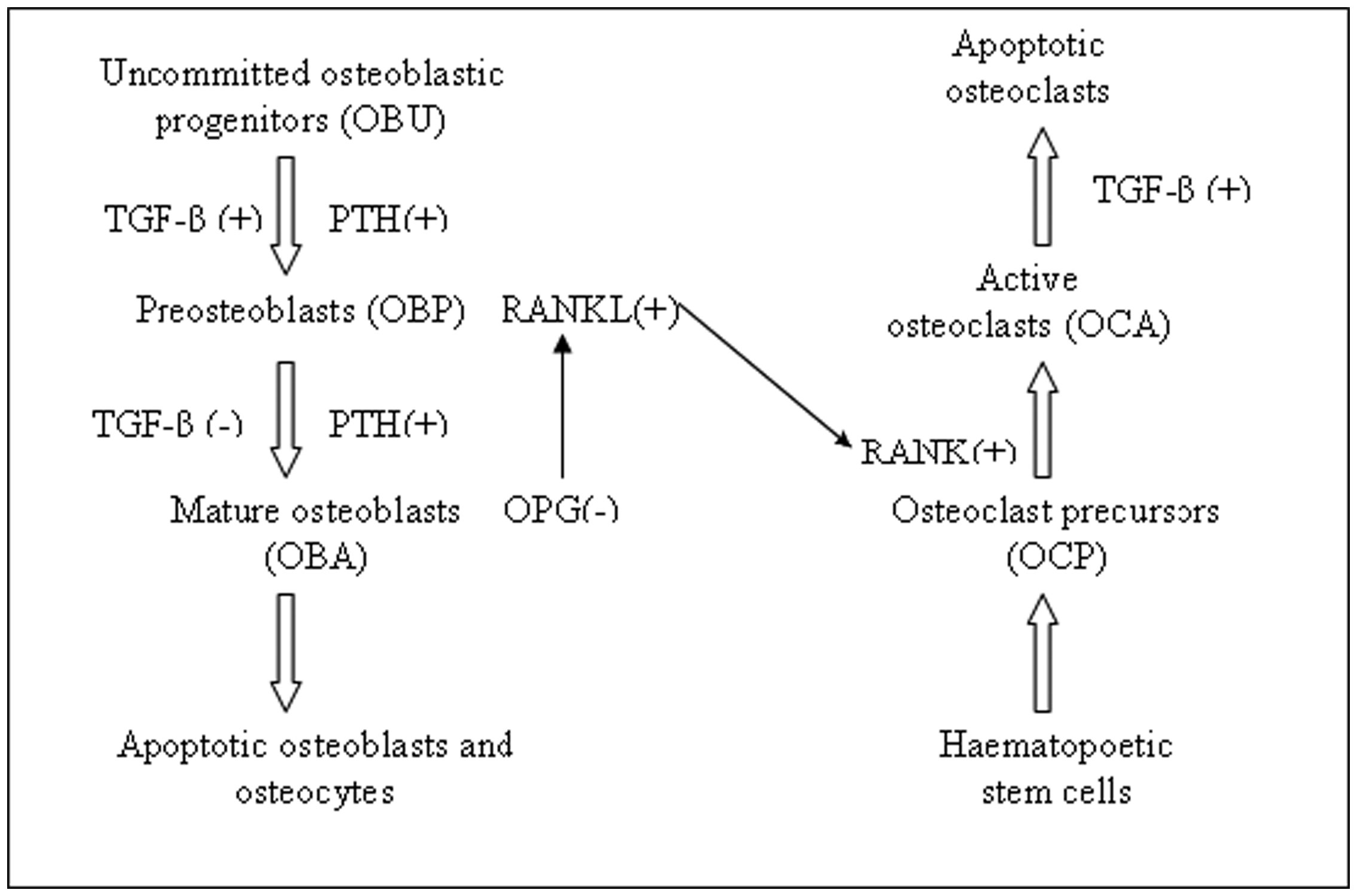
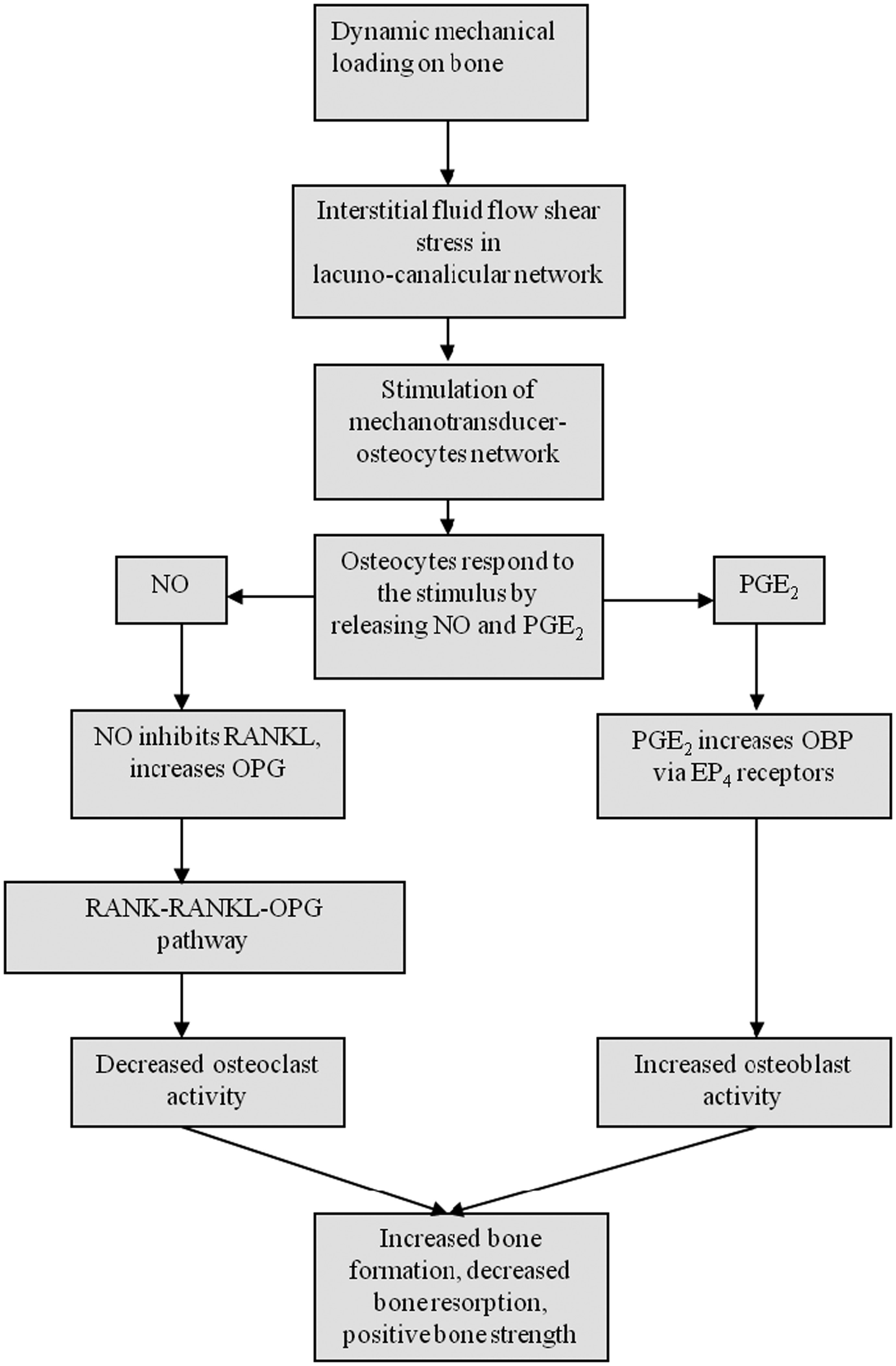
Figures(2)

Chao Hu, Qing-Hua Qin. Bone remodeling and biological effects of mechanical stimulus[J]. AIMS Bioengineering, 2020, 7(1): 12-28. doi: 10.3934/bioeng.2020002







 DownLoad:
DownLoad: