Citation: Frank B Dazzo, Youssef G Yanni, Ashley Jones, Abdelgawad Y Elsadany. CMEIAS bioimage informatics that define the landscape ecology of immature microbial biofilms developed on plant rhizoplane surfaces[J]. AIMS Bioengineering, 2015, 2(4): 469-486. doi: 10.3934/bioeng.2015.4.469

| [1] | Dazzo FB, Gantner S (2012) Rhizosphere. In: Schmidt T, Schaechter M, Topics in Ecological and Environmental Microbiology, Massachusetts, USA: Elsevier Press, 467-479. |
| [2] | Dazzo FB, McGarrell D (2015) CMEIAS Center for Microbial Ecology Image Analysis System. Available from: http://cme.msu.edu/cmeias/. |
| [3] | Dazzo FB (2006) COST 631: Visualization of the rhizoplane microflora by computer-assisted microscopy and spatial analysis by CMEIAS image analysis. In: Luster J., Finlay R., Handbook of Methods Used in Rhizosphere Research, Section 4.1, Birmensdorf, Swiss Federal Research Institute. |
| [4] | Dazzo FB, Mateos P, Orgambide G, et al. (1993) The infection process in the Rhizobium-legume symbiosis and visualization of rhizoplane microorganisms by laser scanning confocal microscopy. In: Guerrero R., Pedros-Alio C., Trends in Microbial Ecology, Barcelona, Spain: Spanish Society for Microbiology, 259-262. |
| [5] |
Folland I, Trione D, Dazzo FB (2014) Accuracy of biovolume formulas for CMEIAS computer-assisted microscopy and body size analysis of morphologically diverse microbial populations and communities. Microbial Ecology 68: 596-610. doi: 10.1007/s00248-014-0410-9

|
| [6] | Loferer-Krobacher M, Klima J, Psenner R (1998) Determination of bacterial cell dry mass by transmission electron microscopy and densitometric image analysis. Appl Environ Microbiol 64: 688-694. |
| [7] | Dazzo FB, Joseph AR, Gomaa A, et al. (2003) Quantitative indices for the autecological biogeography of a Rhizobium endophyte of rice at macro and micro spatial scales. Symbiosis 35: 147-158. |
| [8] | Dazzo FB, Schmid M, Hartmann A (2007) Immunofluorescence microscopy and fluorescence in situ hybridization combined with CMEIAS and other image analysis tools for soil- and plant-associated microbial autecology. In: Garland J, Hurst C, Lipson D, et al., Manual of Environmental Microbiology, Vol 59, 3rd Ed, Washington DC, USA: American Society for Microbiology Press, 712-733. |
| [9] | Yanni YG, Rizk RY, Corish V, et al. (1997) Natural endophytic association between Rhizobium leguminosarum bv. trifolii and rice roots and assessment of its potential to promote rice growth. Plant and Soil 194: 99-114. |
| [10] | Yanni YG, Rizk RY, Abd El-Fattah FK, et al. (2001) The beneficial plant growth-promoting association of Rhizobium leguminosarum bv. trifolii with rice roots. Austr J Plant Physiol 28: 845-870. |
| [11] | Yanni YG, Dazzo FB (2010) Enhancement of rice production using endophytic strains of Rhizobium leguminosarum bv. trifolii in extensive field inoculation trials within the Egypt Nile delta. Plant and Soil 336: 129-142. |
| [12] |
Liu J, Dazzo FB, Glagoleva O, et al. (2001) CMEIAS: a computer-aided system for the image analysis of bacterial morphotypes in microbial communities. Microbial Ecology 41: 173-194. doi: 10.1007/s002480000004
|
| [13] | Gross, CA, Reddy C, Dazzo FB (2010) CMEIAS color segmentation: an improved computing technology to process color images for quantitative microbial ecology studies at single-cell resolution. Microbial Ecology 59: 400-414. |
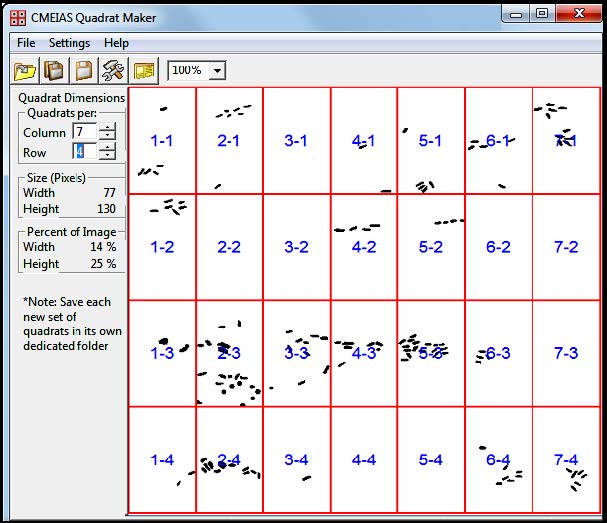
| [14] | Dazzo F, Gross C (2012) CMEIAS Quadrat Maker: a digital software tool to optimize grid dimensions and produce quadrat images for landscape ecology spatial analysis. J Ecosystem and Ecography 3: 4 (136). |
| [15] | Ji Z, Card K, Dazzo FB (2015) CMEIAS JFrad: a new computing toolkit to discriminate the fractal geometry of landscape architecture and spatial patterns of individual cells in microbial biofilms. Microbial Ecology 69: 710-720. |
| [16] | Dazzo FB, Liu J, Gross C, et al. CMEIAS v3.10: Advanced Computational Tools of Image Analysis Software Designed to Strengthen Microscopy-Based Approaches for Understanding Microbial Ecology. All-Investigator Meeting Kellogg Biological Station Long-Term Ecological Research Mtg, 2015. Available from: http://lter.kbs.msu.edu/abstracts/493. |
| [17] | Towner H EcoStat: an Ecological Analysis Program, version 1.03. Exeter Software, Setauket, New York. 1999 available from: http://www.exetersoftware.com/cat/Trinity/ecostat.html. |
| [18] | Krebs C Ecological Methodology, version 7.2, Exeter Software, Setauket, NY, 2011, available from:http://www.exetersoftware.com/cat/ecometh/ecomethodology.html. |
| [19] | Hammer O, Harper DAT, Ryan PD (2001) PAST: Paleontological statistics software package for education and data analysis. Palaeontol Electron 4: 1-9. |
| [20] | Roberts A, Withers P. StatistiXL, Version 1.10. Broadway-Nedlands, Kalamunda, Australia, 2012, available from: http://www.statistixl.com/default.aspx. |
| [21] | Robertson GP GS+ Geostatistics, Version 9.0 Gamma Design Software, Plainwell, MI USA. 2004, Available from: http://www.gammadesign.com/default.aspx. |
| [22] | Paul EA, Harris D, Klug MJ, et al. (1999) The determination of microbial biomass. In: Robertson GP, Coleman DC, Bledsoe CS, et al., Standard Soil Methods for Long-Term Ecological Research. New York, USA: Oxford University Press, 291-317. |
| [23] |
Dazzo FB, Klemmer KJ, Chandler R, et al. (2013) In situ ecophysiology of microbial biofilm communities analyzed by CMEIAS computer-assisted microscopy at single-cell resolution. Diversity 5: 426-460. doi: 10.3390/d5030426
|
| [24] | Petchey OI, Long ZT, Morin PJ (2007) The consequences of body size in model microbial ecosystems. In: Hildrew A, Raffelli D, Edmonds-Brown R, Body Size: the Structure and Function of Aquatic Ecosystems. Cambridge UK: Cambridge University Press, 245-265. |
| [25] | Magurran AE (2004) Measuring Biological Diversity. Malden Massachusetts, USA: Blackwell. |
| [26] | Gonzalez JM, Sherr EB, Sherr BF (1990) Size-selective grazing on bacteria by natural assemblages of estuarine flagellates and ciliates. Appl Environ Microbiol 56: 583-589. |
| [27] |
Fukuda M, Matsuyama J, Katano T et al. (2006) Assessing primary and bacterial production rates in biofilms on pebbles in Ishite Stream, Japan. Microbial Ecology 52: 1-9. doi: 10.1007/s00248-006-9114-0
|
| [28] | Legendre P, Legendre L (2012) Numerical Ecology, 3rd Ed., Amsterdam The Netherlands: Elsevier. |
| [29] | Dazzo FB, Ganesan I, Yanni YG (2014) Spatial ecology of rhizobacterial colonization on roots analyzed by CMEIAS computer-assisted microscopy at single-cell resolution. In: Singh DP, Singh H, Trends in Soil Microbial Ecology, Chapter. 4, Houston USA: Studium Press, 67-92. |
| [30] |
Gantner S, Schmid M, Dürr C, et al. (2006). In situ spatial scale of calling distances and population density-independent N-Acylhomoserine lactone mediated communication by rhizobacteria colonized on plant roots. FEMS Microbiol Ecol 56: 188-194. doi: 10.1111/j.1574-6941.2005.00037.x
|
| [31] |
Dazzo FB (2012) CMEIAS-aided microscopy of the spatial ecology of individual bacterial interactions involving cell-to-cell communication within biofilms. Sensors 12: 7047-7062. doi: 10.3390/s120607047
|
| [32] | Perry J (1998) Measures of spatial patterns for counts. Ecology 79: 1008-1017. |
| [33] | Ripley BD (1979) Tests of “randomness” for spatial point patterns. J Roy Statist Soc B 41: 368-374. |
| [34] | Hammer Ø (2009) New methods for the statistical detection of point alignments. Computers and Geosciences 35: 659-666. |
| [35] |
Robertson GP (1987) Geostatistics in ecology: interpolating with known variance. Ecology 68: 744-748. doi: 10.2307/1938482
|
| [36] | Ettema CH, Wardle DH (2002) Spatial soil ecology. Trends in Ecology and Evolution 17: 177-183. |
| [37] |
Zahid W, Ganczarczyk J (1994) A technique for a characterization of RBC biofilm surface. Water Res 28: 2229-2231. doi: 10.1016/0043-1354(94)90036-1
|
| [38] | Hermanowicz S, Schindler U, Wilderer P (1995) Fractal structure of biofilms: new tools for investigation of morphology. Water Sci Technol 32: 99-105. |
| [39] |
Yang X, Beyenal H, Harkin G, et al. (2000) Quantifying biofilm structure using image analysis. J Microbiol Methods 39: 109-119. doi: 10.1016/S0167-7012(99)00097-4
|
| [40] | Ritchie ME (2010) Scale, Heterogeneity, and the Structure and Diversity of Ecological Communities. Princeton, NJ USA: Princeton University Press. |
| [41] | Sugihara G, May RM (1990) Applications of fractals in ecology. TREE 5: 79-86. |
| [42] | West G, Brown J, Enquist B (1999) The fourth dimension of life: fractal geometry and allometric scaling of organisms. Science 284: 1677-1679. |
| [43] | Dale MRT, Fortin MJ (2014) Spatial Analysis: a Guide for Ecologists. 2nd Ed. Cambridge UK: Cambridge University Press. |
| [44] |
Hill MO (1973) The intensity of spatial pattern in plant communities. J Ecol 61: 225-235. doi: 10.2307/2258930
|
Figures(13) / Tables(2)

Frank B Dazzo, Youssef G Yanni, Ashley Jones, Abdelgawad Y Elsadany. CMEIAS bioimage informatics that define the landscape ecology of immature microbial biofilms developed on plant rhizoplane surfaces[J]. AIMS Bioengineering, 2015, 2(4): 469-486. doi: 10.3934/bioeng.2015.4.469







 DownLoad:
DownLoad: