Citation: Mario Jolicoeur. Modeling cell behavior: moving beyond intuition[J]. AIMS Bioengineering, 2014, 1(1): 1-12. doi: 10.3934/bioeng.2014.1.1

| [1] | Pavlou AK, Belsey MJ (2008) The therapeutic antibodies market to 2008. Eur J Pharm Biopharm 59: 389-396. |
| [2] |
Wurm F (2004) Production of recombinant protein therapeutics in cultivated mammalian cells. Nat Biotechnol 22: 1393-1398. doi: 10.1038/nbt1026

|
| [3] |
Li F, Vijayasankaran N, Shen A, et al. (2010) Cell culture processes for monoclonal antibody production. mAbs J 2: 466-479. doi: 10.4161/mabs.2.5.12720
|
| [4] | Pirt SJ (1975) Principles of microbe and cell cultivation. Blackwell Scientific Publications. |
| [5] | Bailey JE, Ollis DF (1986) Biochemical Engineering Fundamentals, 2 Eds., New York: McGraw-Hill, 984. |
| [6] |
Monod J (1949) The Growth of Bacterial Cultures. Ann Rev Microbiol 3: 371. doi: 10.1146/annurev.mi.03.100149.002103
|
| [7] |
Lamboursain L, Jolicoeur M (2005) Critical influence of Eschscholtzia californica cells nutritional state on secondary metabolite production. Biotechnol Bioeng 91: 827-837. doi: 10.1002/bit.20553
|
| [8] | Cloutier M, Bouchard-Marchand É, Perrier M, et al. (2007) A predictive nutritional model for plant cells and hairy roots. Biotechnol Bioeng 99: 189-200. |
| [9] | Stephanopoulos G, Aristodou, Nielsen J (1998) Metabolic Engineering. Principles and Methodologies, San Diego : Academic Press, 698. |
| [10] | Palsson BO (2011) Systems Biology. Simulation of dynamic network states, Cambridge UK: Cambridge University Press, 317. |
| [11] | Fell D (1997) Understanding the Control of Metabolism. In Frontiers in metabolism. London UK: Portland Press, 301. |
| [12] |
Bordbar A, Monk JM, King ZA, et al. (2014) Constraint-based models predict metabolic and associated cellular functions. Nat Rev Gen 15: 107-120. doi: 10.1038/nrg3643
|
| [13] |
Chong WPK, Thng SH, Hiu AP, et al. (2012) LC-MS-Based Metabolic Characterization of High Monoclonal Antibody-Producing Chinese Hamster Ovary Cells. Biotechnol Bioeng 109:3103-3111. doi: 10.1002/bit.24580
|
| [14] |
Dunn WB, Broadhurst DI, Atherton HJ, et al. (2011) Systems level studies of mammalian metabolomes: the roles of mass spectrometry and nuclear magnetic resonance spectroscopy. Chem Soc Rev 40: 387-426. doi: 10.1039/B906712B
|
| [15] | Ben-Tchavtchavadze M, Perrier M, Jolicoeur M (2010) A Non-Invasive Technique for the Measurement of the Energetic State of Free-Suspension Mammalian Cells. Biotechnol Prog 26: 532-541. |
| [16] | Scheer M, Grote A, Chang A, et al. (2011) BRENDA, the enzyme information system. Nucleic Acids Res 39 (Database issue http://www.brenda-enzymes.org/): 670-676. |
| [17] |
Nolan RP, Lee K (2011) Dynamic model of CHO cell metabolism. Metab Eng 13: 108-124. doi: 10.1016/j.ymben.2010.09.003
|
| [18] |
Leduc M, Tikhomiroff C, Cloutier M, et al. (2006) Development of a kinetic metabolic model: application to Catharanthus roseus hairy root. Bioprocess Biosyst Eng 28: 295-313. doi: 10.1007/s00449-005-0034-z
|
| [19] | Ghorbaniaghdam A, Henry O, Jolicoeur M (2012) A kinetic-metabolic model based on cell energetic state: study of CHO cell behavior under Na-butyrate stimulation, Bioprocess Biosyst Eng 36: 469-487. |
| [20] |
Ghorbaniaghdam A, Chen J, Henry O, et al. (2014) Analyzing clonal variation of monoclonal antibody producing CHO cell lines using an in silico metabolomic platform. PLoS One 9:90832. doi: 10.1371/journal.pone.0090832
|
| [21] | Ghorbaniaghdam A, Chen J, Henry O, et al. (2014) An in silico study of the regulation of CHO cells glycolysis. J Theor Biol In press. |
| [22] |
Cloutier M, Chen J, Tagte F, et al. (2009) Kinetic metabolic modelling for the control of plant cells cytoplasmic phosphate. J Theor Biol 259: 118-131. doi: 10.1016/j.jtbi.2009.02.022
|
| [23] |
Cloutier M, Chen J, DeDobbeleer C, et al. (2009) A systems approach to plant bioprocess optimization. Plant Biotechnol J 7: 939-951. doi: 10.1111/j.1467-7652.2009.00455.x
|
| [24] |
Valancin A, Srinivasan B, Rivoal J, et al. (2013) Analyzing the effect of decreasing cytosolic triosephosphate isomerase on Solanum tuberosum hairy root cells using a kinetic-metabolic model. Biotechnol Bioeng 110: 924-935. doi: 10.1002/bit.24747
|
| [25] | Poliquin PO, Chen J, Cloutier M, et al. (2013) Metabolomics and in-silico analysis reveal critical energy deregulations in animal models of Parkinson's disease. PLoS One 8: 69146 |
| [26] | Michal G, Schomburg D (2012) Biochemical Pathways: An atlas of biochemistry and molecular biology, 2 Eds., Wiley, 398. |
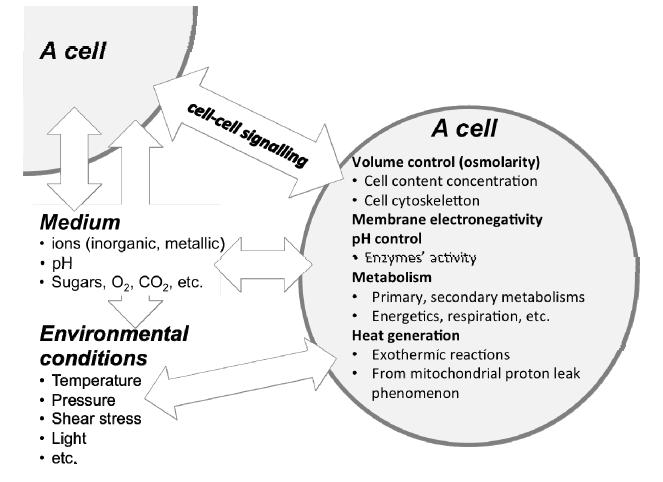

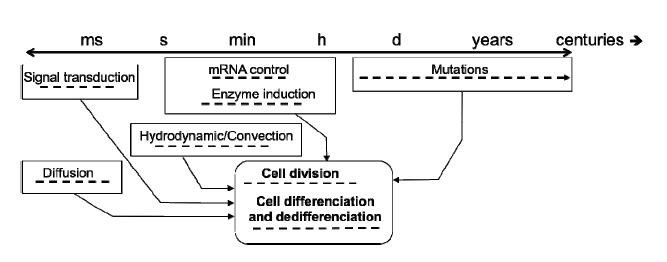
Figures(3)

Mario Jolicoeur. Modeling cell behavior: moving beyond intuition[J]. AIMS Bioengineering, 2014, 1(1): 1-12. doi: 10.3934/bioeng.2014.1.1







 DownLoad:
DownLoad: