Citation: Lukáš Pichl, Taisei Kaizoji. Volatility Analysis of Bitcoin Price Time Series[J]. Quantitative Finance and Economics, 2017, 1(4): 474-485. doi: 10.3934/QFE.2017.4.474

| [1] |
Andersen TG, Bollerslev T, Diebold FX, et al. (2000) Exchange Rate Returns Standardized by Realized Volatility are (Nearly) Gaussian. Multinatl Financ J 4: 159–179. doi: 10.17578/4-3/4-2

|
| [2] |
Andersen TG, Bollerslev T, Diebold FX, et al. (2001) The distribution of realized stock return volatility. J Financ Econ 61: 43–76. doi: 10.1016/S0304-405X(01)00055-1
|
| [3] |
Andersen TG, Bollerslev T, Diebold FX (2007) Roughing it up: including jump components in measuring, modeling and forecasting asset return volatility. Rev Econ Stat 89: 701–720. doi: 10.1162/rest.89.4.701
|
| [4] | Balcilar M, Bouri E, Gupta R, et al. (2017) Can volume predict Bitcoin returns and volatility? A quantiles-based approach. Econ Model 64: 74–81. |
| [5] | Bariviera AF, Basgall MJ, Hasperué W, et al. (2017) Some stylized facts of the Bitcoin market, Physica A: Stat Mechanics Appl 484: 82–90. |
| [6] |
Barndorff-Nielsen OE, Shephard N (2004) Power and Bipower Variation with Stochastic Volatility and Jumps. J Financ Econom 2: 1–37. doi: 10.1093/jjfinec/nbh001
|
| [7] |
Blau BM (2017) Price dynamics and speculative trading in bitcoin. Res Int Bus Financ 41: 493–499. doi: 10.1016/j.ribaf.2017.05.010
|
| [8] | Boudt K, Cornelissen J, Payseur S, et al. (2017) high frequency: Tools for High frequency Data Analysis. R package version 0.5. Available from: https://CRAN.R-project.org/package=highfrequency. |
| [9] | Bouri E, Azzi G, Dyhrberg AH (2017a) On the return-volatility relationship in the Bitcoin market around the price crash of 2013. Econ 11: 1–16. |
| [10] | Bouri E, Molnár P, Azzi G, et al. (2017b) On the hedge and safe haven properties of Bitcoin: Is it really more than a diversifier? Financ Res Letters 20: 192–198. |
| [11] | Bradbury D (2013) The problem with Bitcoin. Comput Fraud Security 2013: 5–8. |
| [12] |
Brandvold M, Molnár P, Vagstad K, et al. (2015) Price discovery on Bitcoin exchanges. J Int Financ Mark, Inst Money 36: 18–35. doi: 10.1016/j.intfin.2015.02.010
|
| [13] | Cheah ET, Fry J (2015) Speculative bubbles in Bitcoin markets? An empirical investigation into the fundamental value of Bitcoin. Econ Lett 130: 32–36. |
| [14] | Dwyer GP (2015) The economics of Bitcoin and similar private digital currencies. J Financ Stab 17: 81–91. |
| [15] | Dyhrberg AH (2016) Hedging capabilities of Bitcoin. Is it the virtual gold? Financ Res Lett 16: 139–144. |
| [16] |
Extance A (2015) Bitcoin and beyond. Nature 526: 21–23. doi: 10.1038/526021a
|
| [17] | Franzke C (2012) Predictability of extreme events in a nonlinear stochastic-dynamical model. Physical Rev E 85. |
| [18] | Fritsch S, Guenther F (2016) neuralnet: Training of Neural Networks. R package version 1.33. Available online: https://CRAN.R-project.org/package=neuralnet. |
| [19] | Hawkes AG, Oakes D (1974) A Cluster Process Representation of a Self-Exciting Process. J Appl Prob 11: 493–503. |
| [20] | Hsieh WW (2009) Machine Learning Methods in the Environmental Sciences, Cambridge University Press, Cambridge, UK. |
| [21] | Katsiampa P (2017) Volatility estimation for Bitcoin: A comparison of GARCH models. Econ Lett 158: 3–6. |
| [22] | Kim YB, Kim JG, Kim W, et al. (2016) Predicting Fluctuations in Cryptocurrency Transactions Based on User Comments and Replies. PLoS ONE 11. |
| [23] | Kristoufek L (2013) BitCoin meets Google Trends and Wikipedia: Quantifying the relationship between phenomena of the Internet era. Scientific rep 3. |
| [24] | Kristoufek L (2015) What Are the Main Drivers of the Bitcoin Price? Evidence from Wavelet Coherence Analysis. PLoS One 10. |
| [25] |
Urquhart A (2016) The inefficiency of Bitcoin. Econ Lett 148: 80–82. doi: 10.1016/j.econlet.2016.09.019
|
| [26] |
Urquhart A (2017) Price clustering in Bitcoin. Econ Lett 159: 145–148. doi: 10.1016/j.econlet.2017.07.035
|
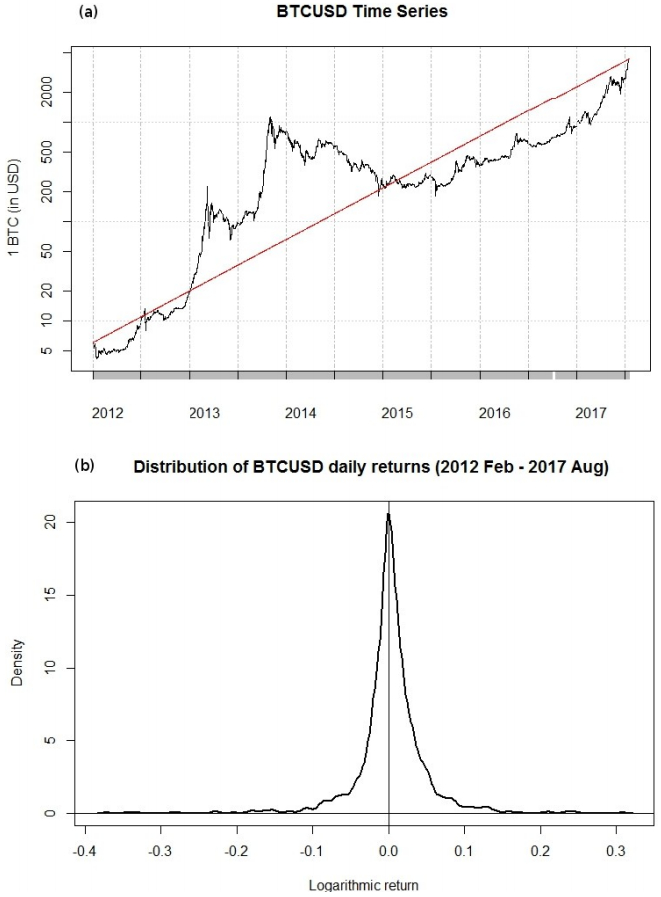
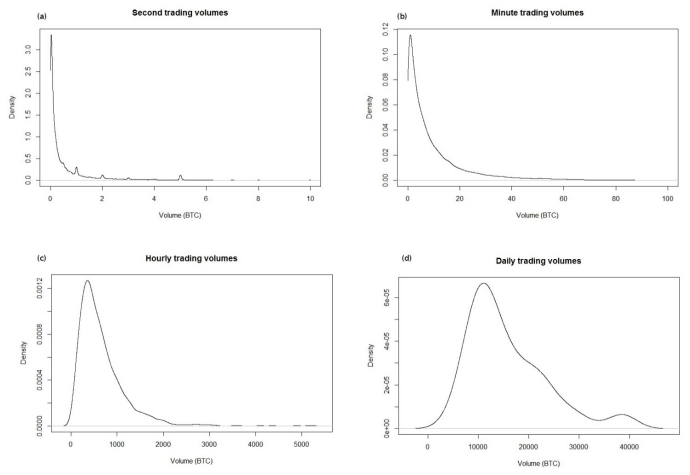
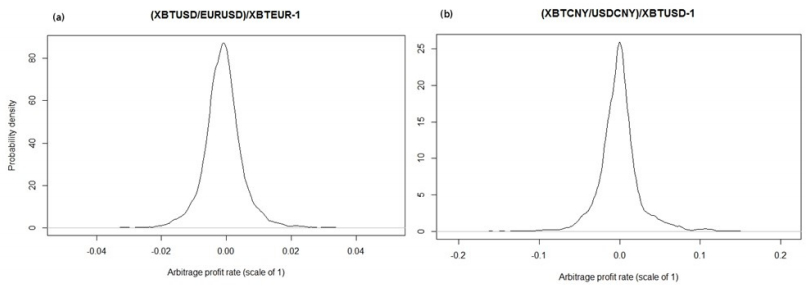
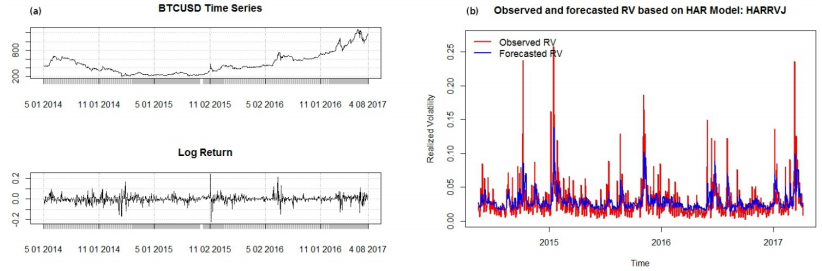
Figures(5) / Tables(1)

Lukáš Pichl, Taisei Kaizoji. Volatility Analysis of Bitcoin Price Time Series[J]. Quantitative Finance and Economics, 2017, 1(4): 474-485. doi: 10.3934/QFE.2017.4.474







 DownLoad:
DownLoad: