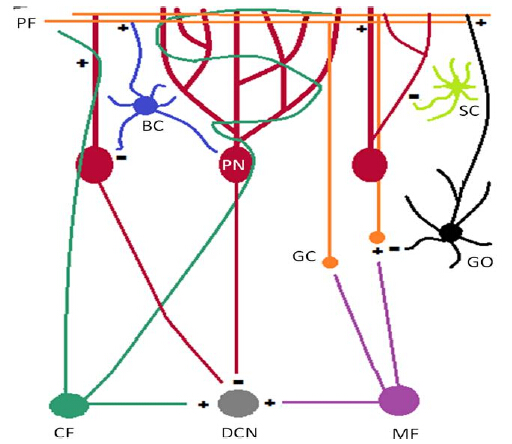
The notion that the cerebellum is a central regulator of motor function is undisputed. There exists, however, considerable literature to document a similarly vital role for the cerebellum in the regulation of various non-motor domains, including emotion. Research from numerous avenues of investigation (i.e., neurophysiological, behavioural, electrophysiological, imagining, lesion, and clinical studies) have documented the importance of the cerebellum, in particular, the vermis, in affective processing that appears preserved across species. The cerebellum possesses a distinct laminar arrangement and highly organized neuronal circuitry. Moreover, the cerebellum forms reciprocal connections with several brain regions implicated in diverse functional domains, including motor, sensory, and emotional processing. It has been argued that these unique neuroanatomical features afford the cerebellum with the capacity to integrate information about an organism, its environment, and its place within the environment such that it can respond in an appropriate, coordinated fashion, with such theories extending to the regulation of emotion. This review puts our current understanding of the cerebellum and its role in behaviour in historical perspective, presents an overview of the neuroanatomical and functional organization of the cerebellum, and reviews the literature describing the involvement of the cerebellum in emotional regulation in both humans and non-human animals. In summary, this review discusses the importance of the functional connectivity of the cerebellum with various brain regions in the ability of the cerebellum to effectively regulate emotional behaviour.
Citation: Wanda M. Snow, Brenda M. Stoesz, Judy E. Anderson. The Cerebellum in Emotional Processing: Evidence from Human and Non-Human Animals[J]. AIMS Neuroscience, 2014, 1(1): 96-119. doi: 10.3934/Neuroscience.2014.1.96
The notion that the cerebellum is a central regulator of motor function is undisputed. There exists, however, considerable literature to document a similarly vital role for the cerebellum in the regulation of various non-motor domains, including emotion. Research from numerous avenues of investigation (i.e., neurophysiological, behavioural, electrophysiological, imagining, lesion, and clinical studies) have documented the importance of the cerebellum, in particular, the vermis, in affective processing that appears preserved across species. The cerebellum possesses a distinct laminar arrangement and highly organized neuronal circuitry. Moreover, the cerebellum forms reciprocal connections with several brain regions implicated in diverse functional domains, including motor, sensory, and emotional processing. It has been argued that these unique neuroanatomical features afford the cerebellum with the capacity to integrate information about an organism, its environment, and its place within the environment such that it can respond in an appropriate, coordinated fashion, with such theories extending to the regulation of emotion. This review puts our current understanding of the cerebellum and its role in behaviour in historical perspective, presents an overview of the neuroanatomical and functional organization of the cerebellum, and reviews the literature describing the involvement of the cerebellum in emotional regulation in both humans and non-human animals. In summary, this review discusses the importance of the functional connectivity of the cerebellum with various brain regions in the ability of the cerebellum to effectively regulate emotional behaviour.

| [1] | Andersen BB, Korbo L, Pakkenberg B. (1992) A quantitative study of the human cerebellum with unbiased stereological techniques. J Comp Neurol 326(4): 549-60. |
| [2] | Glickstein M, Doron K. (2008) Cerebellum: Connections and functions. Cerebellum 7(4):589-94. |
| [3] | Gibb R, Kolb B. (1999) A method for vibratome sectioning of golgi-cox stained whole rat brain. J Neurosci Methods 79(1): 1-4. |
| [4] |
Ito M. (2002) Historical review of the significance of the cerebellum and the role of purkinje cells in motor learning. Ann N Y Acad Sci 978: 273-88. doi: 10.1111/j.1749-6632.2002.tb07574.x

|
| [5] | Eccles JC, Ito M, Szentágothai J. (1967) The Cerebellum as a Neuronal Machine. New York: NY Springer-Verlag. |
| [6] | Marr D. (1960) A theory of cerebellar cortex. J Physiol 202(2): 437-70. |
| [7] |
Albus JS. (1971) A theory of cerebellar function. Math Biosci 10: 25-61. doi: 10.1016/0025-5564(71)90051-4
|
| [8] |
Thach WT, Goodkin HP, Keating JG. (1992) The cerebellum and the adaptive coordination of movement. Ann Rev Neurosci 15: 403-42. doi: 10.1146/annurev.ne.15.030192.002155
|
| [9] | Leiner HC, Leiner AL, Dow RS. (1986) Does the cerebellum contribute to mental skills? Behav Neurosci 100(4): 443-54. |
| [10] | Keele SW, Ivry R. (1990) Does the cerebellum provide a common computation for diverse tasks? A timing hypothesis. Ann NY Acad Sci 608: 207-11. |
| [11] |
Petersen SE, Fox PT, Posner MI, et al. (1989) Positron emission tomographic studies of the processing of single words. J Cognitive Neurosci 1: 153-170. doi: 10.1162/jocn.1989.1.2.153
|
| [12] |
Strick PL, Dum RP, Fiez JA. (2009) Cerebellum and nonmotor function. Annu Rev Neurosci 32:413-34. doi: 10.1146/annurev.neuro.31.060407.125606
|
| [13] |
DOW RS. (1961) Some aspects of cerebellar physiology. J Neurosurg 18: 512-30. doi: 10.3171/jns.1961.18.4.0512
|
| [14] | Kandel ER, Schwartz JH, Jessell TM. (2000) Principles of neural science. 4Eds, New York: McGraw-Hill, Health Professions Division, 1414 . |
| [15] |
Berntson GG, Torello MW. (1982) The paleocerebellum and the integration of behavioural function. Physiol Psychol 10: 2-12. doi: 10.3758/BF03327003
|
| [16] |
Middleton FA, Strick PL. (1997) Cerebellar output channels. Int Rev Neurobiol 41: 61-82. doi: 10.1016/S0074-7742(08)60347-5
|
| [17] | Hashimoto M, Hibi M. (2012) Development and evolution of cerebellar neural circuits. Dev Growth Differ 54(3): 373-89. |
| [18] | Ito M. (2006) Cerebellar circuitry as a neuronal machine. Prog Neurobiol 78(3-5): 272-303. |
| [19] | Eccles JC. (1967) Circuits in the cerebellar control of movement. Proc Natl Acad Sci USA58(1): 336-43. |
| [20] | Akshoomoff NA, Courchesne E. (1992) A new role for the cerebellum in cognitive operations. Behav Neurosci 106(5): 731-8. |
| [21] | Decety J, Sjoholm H, Ryding E, Stenberg G, Ingvar DH. (1990) The cerebellum participates in mental activity: Tomographic measurements of regional cerebral blood flow. Brain Res 535(2):313-7. |
| [22] | Ryding E, Decety J, Sjoholm H, et al. (1993) Motor imagery activates the cerebellum regionally. A SPECT rCBF study with 99mTc-HMPAO. Brain Res Cogn Brain Res 1(2): 94-9. |
| [23] | Kim SG, Ugurbil K, Strick PL. (1994) Activation of a cerebellar output nucleus during cognitive processing. Science 265(5174): 949-51. |
| [24] |
Schmahmann JD, Sherman JC. (1998) The cerebellar cognitive affective syndrome. Brain 121:561-79. doi: 10.1093/brain/121.4.561
|
| [25] | Exner C, Weniger G, Irle E. (2004) Cerebellar lesions in the PICA but not SCA territory impair cognition. Neurology 63(11): 2132-5. |
| [26] | Marien P, de Smet HJ, Wijgerde E, et al. (2013) Posterior fossa syndrome in adults: A new case and comprehensive survey of the literature. Cortex 49(1): 284-300. |
| [27] | Kuper M, Timmann D. (2013) Cerebellar mutism. Brain Lang 127(3): 327-33. |
| [28] | Baillieux H, De Smet HJ, Paquier PF, et al. (2008) Cerebellar neurocognition: Insights into the bottom of the brain. Clin Neurol Neurosurg 110(8): 763-73. |
| [29] | Steinlin M, Imfeld S, Zulauf P, et al. (2003) Neuropsychological long-term sequelae after posterior fossa tumour resection during childhood. Brain 126(Pt 9): 1998-2008. |
| [30] |
Levisohn L, Cronin-Golomb A, Schmahmann JD. (2000) Neuropsychological consequences of cerebellar tumour resection in children: Cerebellar cognitive affective syndrome in a paediatric population. Brain 123: 1041-50. doi: 10.1093/brain/123.5.1041
|
| [31] | de Smet HJ, Baillieux H, Wackenier P, et al. (2009) Long-term cognitive deficits following posterior fossa tumor resection: A neuropsychological and functional neuroimaging follow-up study. Neuropsychology 23(6): 694-704. |
| [32] | Pollack IF, Polinko P, Albright AL, et al. (1995) Mutism and pseudobulbar symptoms after resection of posterior fossa tumors in children: Incidence and pathophysiology. Neurosurgery37(5): 885-93. |
| [33] | Ozimek A, Richter S, Hein-Kropp C, et al. (2004) Cerebellar mutism––report of four cases. J Neurol 251(8): 963-72. |
| [34] | Daniels SR, Moores LE, DiFazio MP. (2005) Visual disturbance associated with postoperative cerebellar mutism. Pediatr Neurol 32(2): 127-30. |
| [35] | Riva D. (1998) The cerebellar contribution to language and sequential functions: Evidence from a child with cerebellitis. Cortex 34(2): 279-87. |
| [36] | Drost G, Verrips A, Thijssen HO, et al. (2000) Cerebellar involvement as a rare complication of pneumococcal meningitis. Neuropediatrics 31(2): 97-9. |
| [37] | Fujisawa H, Yonaha H, Okumoto K, et al. (2005) Mutism after evacuation of acute subdural hematoma of the posterior fossa. Childs Nerv Syst 21(3): 234-6. |
| [38] | Ersahin Y, Mutluer S, Saydam S, Barcin E. (1997) Cerebellar mutism: Report of two unusual cases and review of the literature. Clin Neurol Neurosurg 99(2): 130-4. |
| [39] | Al-Anazi A, Hassounah M, Sheikh B, et al. (2001) Cerebellar mutism caused by arteriovenous malformation of the vermis. Br J Neurosurg 15(1): 47-50. |
| [40] | Dubey A, Sung WS, Shaya M, et al. (2009) Complications of posterior cranial fossa surgery––an institutional experience of 500 patients. Surg Neurol 72(4): 369-75. |
| [41] | Afshar-Oromieh A, Linhart H, Podlesek D, et al. (2010) Postoperative cerebellar mutism in adult patients with lhermitte-duclos disease. Neurosurg Rev 33(4): 401-8. |
| [42] | Coplin WM, Kim DK, Kliot M, et al. (1997) Mutism in an adult following hypertensive cerebellar hemorrhage: Nosological discussion and illustrative case. Brain Lang 59(3): 473-93. |
| [43] | Moore MT. (1969) Progressive akinetic mutism in cerebellar hemangioblastoma with "normal-pressure hydrocephalus". Neurology 19(1): 32-6. |
| [44] | Caner H, Altinors N, Benli S, et al. (1999) Akinetic mutism after fourth ventricle choroid plexus papilloma: Treatment with a dopamine agonist. Surg Neurol 51(2): 181-4. |
| [45] | Idiaquez J, Fadic R, Mathias CJ. (2001) Transient orthostatic hypertension after partial cerebellar resection. Clin Auton Res 21(1): 57-9. |
| [46] | De Smet HJ, Marien P. (2012) Posterior fossa syndrome in an adult patient following surgical evacuation of an intracerebellar haematoma. Cerebellum 11(2): 587-92. |
| [47] | Dunwoody GW, Alsagoff ZS, Yuan SY. (1997) Cerebellar mutism with subsequent dysarthria in an adult: Case report. Br J Neurosurg 11(2): 161-3. |
| [48] | Marien P, Verslegers L, Moens M, et al. (2013) Posterior fossa syndrome after cerebellar stroke. Cerebellum 12(5): 686-91. |
| [49] | Sacchetti B, Scelfo B, Strata P. (2005) The cerebellum: Synaptic changes and fear conditioning. Neuroscientist 11(3): 217-27. |
| [50] | LeDoux JE. (1994) Emotion, memory and the brain. Sci Am 270(6): 50-7. |
| [51] | Maschke M, Schugens M, Kindsvater K, et al. (2002) Fear conditioned changes of heart rate in patients with medial cerebellar lesions. J Neurol Neurosurg Psychiatry 72(1): 116-8. |
| [52] | Maschke M, Drepper J, Kindsvater K, et al. (2000) Fear conditioned potentiation of the acoustic blink reflex in patients with cerebellar lesions. J Neurol Neurosurg Psychiatry 68(3):358-64. |
| [53] | Turner BM, Paradiso S, Marvel CL, et al. (2007) The cerebellum and emotional experience. Neuropsychologia 45(6): 1331-41. |
| [54] | Nashold BS, Wilson WP, Slaughter DG. (1969) Sensations evoked by stimulation in the midbrain of man. J Neurosurg 30(1): 14-24. |
| [55] | Damasio AR, Grabowski TJ, Bechara A, et al. (2000) Subcortical and cortical brain activity during the feeling of self-generated emotions. Nat Neurosci 3(10): 1049-56. |
| [56] | Ploghaus A, Tracey I, Clare S, et al. (2000) Learning about pain: The neural substrate of the prediction error for aversive events. Proc Natl Acad Sci USA 97(16): 9281-6. |
| [57] | Ploghaus A, Tracey I, Gati JS, et al. (1999) Dissociating pain from its anticipation in the human brain. Science 284(5422): 1979-81. |
| [58] | Singer T, Seymour B, O'Doherty J, et al. (2004) Empathy for pain involves the affective but not sensory components of pain. Science 303(5661): 1157-62. |
| [59] | Fischer H, Andersson JL, Furmark T, et al. (2000) Fear conditioning and brain activity: A positron emission tomography study in humans. Behav Neurosci 114(4): 671-80. |
| [60] | Frings M, Maschke M, Erichsen M, et al. (2002) Involvement of the human cerebellum in fear-conditioned potentiation of the acoustic startle response: A PET study. Neuroreport 13(10):1275-8. |
| [61] | Timmann D, Drepper J, Frings M, et al. (2010) The human cerebellum contributes to motor, emotional and cognitive associative learning. A review. Cortex 46(7): 845-57. |
| [62] | Baumann O, Mattingley JB. (2012) Functional topography of primary emotion processing in the human cerebellum. Neuroimage 61(4): 805-11. |
| [63] |
Schienle A, Scharmuller W. (2013) Cerebellar activity and connectivity during the experience of disgust and happiness. Neuroscience 246: 375-81. doi: 10.1016/j.neuroscience.2013.04.048
|
| [64] | Schienle A, Stark R, Walter B, et al. (2002) The insula is not specifically involved in disgust processing: An fMRI study. Neuroreport 13(16): 2023-6. |
| [65] | Stoodley CJ, Schmahmann JD. (2009) Functional topography in the human cerebellum: A meta-analysis of neuroimaging studies. Neuroimage 44(2): 489-501. |
| [66] | Heilman KM, Gilmore RL. (1998) Cortical influences in emotion. J Clin Neurophysiol 15(5):409-23. |
| [67] | Fusar-Poli P, Placentino A, Carletti F, et al. (2009) Functional atlas of emotional faces processing: A voxel-based meta-analysis of 105 functional magnetic resonance imaging studies. J Psychiatry Neurosci 34(6): 418-32. |
| [68] | Vuilleumier P, Pourtois G. (2007) Distributed and interactive brain mechanisms during emotion face perception: Evidence from functional neuroimaging. Neuropsychologia 45(1): 174-94. |
| [69] | Schutter DJ, Enter D, Hoppenbrouwers SS. (2009) High-frequency repetitive transcranial magnetic stimulation to the cerebellum and implicit processing of happy facial expressions. J Psychiatry Neurosci 34(1): 60-5. |
| [70] | Ferrucci R, Giannicola G, Rosa M, et al. (2012) Cerebellum and processing of negative facial emotions: Cerebellar transcranial DC stimulation specifically enhances the emotional recognition of facial anger and sadness. Cogn Emot 26(5):786-99. |
| [71] | Adamaszek M, D'Agata F, Kirkby KC, et al. (2014) Impairment of emotional facial expression and prosody discrimination due to ischemic cerebellar lesions. Cerebellum 13(3):338-45. |
| [72] | Furl N, van Rijsbergen NJ, Kiebel SJ, et al. (2010) Modulation of perception and brain activity by predictable trajectories of facial expressions. Cereb Cortex 20(3): 694-703. |
| [73] | Kilts CD, Egan G, Gideon DA, et al. (2003) Dissociable neural pathways are involved in the recognition of emotion in static and dynamic facial expressions. Neuroimage 18(1): 156-68. |
| [74] | Villanueva R. (2012) The cerebellum and neuropsychiatric disorders. Psychiatry Res 198(3):527-32. |
| [75] | Loeber RT, Cintron CM, Yurgelun-Todd DA. (2001) Morphometry of individual cerebellar lobules in schizophrenia. Am J Psychiatry 158(6): 952-4. |
| [76] | Okugawa G, Sedvall GC, Agartz I. (2003) Smaller cerebellar vermis but not hemisphere volumes in patients with chronic schizophrenia. Am J Psychiatry 160(9): 1614-7. |
| [77] | Jacobsen LK, Giedd JN, Berquin PC, et al. (1997) Quantitative morphology of the cerebellum and fourth ventricle in childhood-onset schizophrenia. Am J Psychiatry 154(12): 1663-9. |
| [78] | Nasrallah HA, Schwarzkopf SB, Olson SC, et al. (1991) Perinatal brain injury and cerebellar vermal lobules I-X in schizophrenia. Biol Psychiatry 29(6): 567-74. |
| [79] | Ichimiya T, Okubo Y, Suhara T, et al. (2001) Reduced volume of the cerebellar vermis in neuroleptic-naive schizophrenia. Biol Psychiatry 49(1): 20-7. |
| [80] | Staal WG, Hulshoff Pol HE, Schnack HG, et al. (2000) Structural brain abnormalities in patients with schizophrenia and their healthy siblings. Am J Psychiatry 157(3): 416-21. |
| [81] | Aylward EH, Reiss A, Barta PE, et al. (1994) Magnetic resonance imaging measurement of posterior fossa structures in schizophrenia. Am J Psychiatry 151(10): 1448-52. |
| [82] | Heath RG. (1977) Modulation of emotion with a brain pacemaker. Treatment for intractable psychiatric illness. J Nerv Ment Dis 165(5): 300-17. |
| [83] | Demirtas-Tatlidede A, Freitas C, Cromer JR, et al. (2010) Safety and proof of principle study of cerebellar vermal theta burst stimulation in refractory schizophrenia. Schizophr Res 124(1-3):91-100. |
| [84] | Manu P, Sarpal D, Muir O, et al. (2011) When can patients with potentially life-threatening adverse effects be rechallenged with clozapine? A systematic review of the published literature. Schizophr Res 134(2-3): 180-6. |
| [85] | Soares JC, Mann JJ. (1997) The anatomy of mood disorders––review of structural neuroimaging studies. Biol Psychiatry 41(1): 86-106. |
| [86] | Fitzgerald PB, Laird AR, Maller J, et al. (2008) A meta-analytic study of changes in brain activation in depression. Hum Brain Mapp 29(6): 683-95. |
| [87] | Leroi I, O'Hearn E, Marsh L, Lyketsos CG, et al. (2002) Psychopathology in patients with degenerative cerebellar diseases: A comparison to huntington's disease. Am J Psychiatry 159(8):1306-14. |
| [88] | Alalade E, Denny K, Potter G. (2011) Altered cerebellar-cerebral functional connectivity in geriatric depression. PLoS One 6(5): e20035. |
| [89] | Kanner L. (1943) Autistic disturbances of affective contact. Nervous Child 2: 217-50. |
| [90] | Senju A, Johnson MH. (2009) Atypical eye contact in autism: Models, mechanisms and development. Neurosci Biobehav Rev 33(8): 1204-14. |
| [91] | Ospina MB, Krebs Seida J, Clark B, et al. (2008) Behavioural and developmental interventions for autism spectrum disorder: A clinical systematic review. PLoS One 3(11): e3755. |
| [92] | Hughes JR. (2009) Update on autism: A review of 1300 reports published in 2008. Epilepsy Behav 16(4): 569-89. |
| [93] | Muratori F, Maestro S. (2007) Autism as a downstream effect of primary difficulties in intersubjectivity interacting with abnormal development of brain connectivity. Inter J Dialog Science 2007(2): 93-118. |
| [94] | Bailey A, Luthert P, Dean A, et al. (1998) A clinicopathological study of autism. Brain 121 ( Pt 5)(Pt 5): 889-905. |
| [95] | Bauman M, Kemper TL. (1985) Histoanatomic observations of the brain in early infantile autism. Neurology 35(6): 866-74. |
| [96] | Courchesne E, Yeung-Courchesne R, Press GA, et al. (1988) Hypoplasia of cerebellar vermal lobules VI and VII in autism. N Engl J Med 318(21): 1349-54. |
| [97] | Courchesne E, Hesselink JR, Jernigan TL, et al. (1987) Abnormal neuroanatomy in a nonretarded person with autism. unusual findings with magnetic resonance imaging. Arch Neurol 44(3):335-41. |
| [98] | Hashimoto T, Tayama M, Murakawa K, et al. (1995) Development of the brainstem and cerebellum in autistic patients. J Autism Dev Disord 25(1):1-18. |
| [99] | Ingram JL, Peckham SM, Tisdale B, et al. (2000) Prenatal exposure of rats to valproic acid reproduces the cerebellar anomalies associated with autism. Neurotoxicol Teratol 22(3): 319-24. |
| [100] | Pierce K, Courchesne E. (2001) Evidence for a cerebellar role in reduced exploration and stereotyped behavior in autism. Biol Psychiatry 49(8): 655-64. |
| [101] | Sparks BF, Friedman SD, Shaw DW, et al. (2002) Brain structural abnormalities in young children with autism spectrum disorder. Neurology 59(2): 184-92. |
| [102] | Allen G, Courchesne E. (2003) Differential effects of developmental cerebellar abnormality on cognitive and motor functions in the cerebellum: An fMRI study of autism. Am J Psychiatry160(2): 262-73. |
| [103] | Waterhouse L, Fein D, Modahl C. (1996) Neurofunctional mechanisms in autism. Psychol Rev103(3): 457-89. |
| [104] | Snow WM, Hartle K, Ivanco TL. (2008) Altered morphology of motor cortex neurons in the VPA rat model of autism. Dev Psychobiol 50(7): 633-9. |
| [105] | Allen G, Buxton RB, Wong EC, et al. (1997) Attentional activation of the cerebellum independent of motor involvement. Science 275(5308): 1940-3. |
| [106] | Lalonde R, Strazielle C. (2003) The effects of cerebellar damage on maze learning in animals. Cerebellum 2(4): 300-9. |
| [107] | Joyal CC, Strazielle C, Lalonde R. (2001) Effects of dentate nucleus lesions on spatial and postural sensorimotor learning in rats. Behav Brain Res 122(2): 131-7. |
| [108] | Joyal CC, Meyer C, Jacquart G, et al. (1996) Effects of midline and lateral cerebellar lesions on motor coordination and spatial orientation. Brain Res 739(1-2): 1-11. |
| [109] | Leggio MG, Molinari M, Neri P, et al. (2000) Representation of actions in rats: The role of cerebellum in learning spatial performances by observation. Proc Natl Acad Sci USA 97(5):2320-5. |
| [110] | Dow RS, Moruzzi G. (1958) The physiology and pathology of the cerebellum. Minneapolis: University of Minnesota Press. |
| [111] | Sprague JM, Chambers WW. (1959) An analysis of cerebellar function in the cat, as revealed by its complete and partial destruction, and its interaction with the cerebral cortex. Archs ital Biol 97:68-88. |
| [112] |
Chambers WW, Sprague JM. (1955) Functional localization in the cerebellum II. Arch Neurol Psychiat (Chic) 74: 653-680. doi: 10.1001/archneurpsyc.1955.02330180071008
|
| [113] | Peters M, Monjan AA. (1971) Behavior after cerebellar lesions in cats and monkeys. Physiol Behav 6(2): 205-6. |
| [114] | Berman AS, Berman D, Prescott JW. (1974) The effect of cerebellar lesions on emotional behavior in the rhesus monkey. In: Cooper IS, Riklan M, Snider RS, editors. Ihe Cerebellum, Epilepsy, and Behavior. New York: Plenum, 1974. |
| [115] | Supple WF, Leaton RN, Fanselow MS. (1987) Effects of cerebellar vermal lesions on species-specific fear responses, neophobia, and taste-aversion learning in rats. Physiol Behav39(5): 579-86. |
| [116] | Barnett SA, Barnett SA. (1975) The rat: a study in behavior. Chicago: University of Chicago. Press. |
| [117] | Supple WF, Leaton RN. (1990) Cerebellar vermis: Essential for classically conditioned bradycardia in the rat. Brain Res 509(1): 17-23. |
| [118] | Supple WF, Kapp BS. (1993) The anterior cerebellar vermis: Essential involvement in classically conditioned bradycardia in the rabbit. J Neurosci 13(9): 3705-11. |
| [119] | Yuzaki M. (2003) The delta2 glutamate receptor: 10 years later. Neurosci Res 46(1): 11-22. |
| [120] | Sacchetti B, Scelfo B, Tempia F, et al. (2004) Long-term synaptic changes induced in the cerebellar cortex by fear conditioning. Neuron 42(6): 973-82. |
| [121] | Saab CY, Willis WD. (2003) The cerebellum: Organization, functions and its role in nociception. Brain Res Brain Res Rev 42(1): 85-95. |
| [122] | Sacchetti B, Baldi E, Lorenzini CA, et al. (2002) Cerebellar role in fear-conditioning consolidation. Proc Natl Acad Sci USA 99(12): 8406-11. |
| [123] | Sacchetti B, Scelfo B, Strata P. (2009) Cerebellum and emotional behavior. Neuroscience 162(3):756-62. |
| [124] | Butler AB, Hodos H. (1996) Comparative vertebrate neuroanatomy. New York: Wiley-Liss. |
| [125] | Rodriguez F, Duran E, Gomez A, et al. (2005) Cognitive and emotional functions of the teleost fish cerebellum. Brain Res Bull 66(4-6): 365-70. |
| [126] |
Yoshida M, Kondo H. (2012) Fear conditioning-related changes in cerebellar purkinje cell activities in goldfish. Behav Brain Funct 8: 52. doi: 10.1186/1744-9081-8-52
|
| [127] | Supple WF, Sebastiani L, Kapp BS. (1993) Purkinje cell responses in the anterior cerebellar vermis during pavlovian fear conditioning in the rabbit. Neuroreport 4(7): 975-8. |
| [128] | Middleton FA, Strick PL. (1994) Anatomical evidence for cerebellar and basal ganglia involvement in higher cognitive function. Science 266(5184): 458-61. |
| [129] | Middleton FA, Strick PL. (2000) Cerebellar projections to the prefrontal cortex of the primate. J Neurosci 21(2):700-12. |
| [130] | Middleton FA, Strick PL. (1998) Cerebellar output: Motor and cognitive channels. Trends Cogn Sci 2(9): 348-54. |
| [131] | Dietrichs E. (1984) Cerebellar autonomic function: Direct hypothalamocerebellar pathway. Science 223(4636): 591-3. |
| [132] | Haines DE, Dietrichs E. (1984) An HRP study of hypothalamo-cerebellar and cerebello-hypothalamic connections in squirrel monkey (saimiri sciureus). J Comp Neurol 229(4):559-75. |
| [133] | Snider RS, Maiti A. (1976) Cerebellar contributions to the papez circuit. J Neurosci Res 2(2):133-46. |
| [134] | Heath RG. (1972) Physiologic basis of emotional expression: Evoked potential and mirror focus studies in rhesus monkeys. Biol Psychiatry 5(1):15-31. |
| [135] | Heath RG, Harper JW. (1974) Ascending projections of the cerebellar fastigial nucleus to the hippocampus, amygdala, and other temporal lobe sites: Evoked potential and histological studies in monkeys and cats. Exp Neurol 45(2): 268-87. |
| [136] | Prescott JW. (1971) Early somatosensory deprivation as ontogenic process in the abnormal development of the brain and behavior. In: Moor-Jankowski E, editor. Basel: Karger. |
| [137] | D'Angelo E, Casali S. (2013) Seeking a unified framework for cerebellar function and dysfunction: From circuit operations to cognition. Front Neural Circuits 6: 116. |
| [138] | Larouche M, Hawkes R. (2006) From clusters to stripes: The developmental origins of adult cerebellar compartmentation. Cerebellum 5(2): 77-88. |
| [139] |
Snow WM, Anderson JE, Fry M. (2014) Regional and genotypic differences in intrinsic electrophysiological properties of cerebellar purkinje neurons from wild-type and dystrophin-deficient mdx mice. Neurobiol Learn Mem 107: 19-31. doi: 10.1016/j.nlm.2013.10.017
|
| [140] | Stoodley CJ, Schmahmann JD. (2010) Evidence for topographic organization in the cerebellum of motor control versus cognitive and affective processing. Cortex 46(7): 831-44. |
| [141] | Schmahmann JD, Caplan D. (2006) Cognition, emotion and the cerebellum. Brain 129(Pt 2):290-2. |
| [142] | Leiner HC, Leiner AL, Dow RS. (1989) Reappraising the cerebellum: What does the hindbrain contribute to the forebrain? Behav Neurosci 103(5): 998-1008. |
| [143] | Schmahmann JD. (2010) The role of the cerebellum in cognition and emotion: Personal reflections since 1982 on the dysmetria of thought hypothesis, and its historical evolution from theory to therapy. Neuropsychol Rev 20(3): 236-60. |
| [144] | Nudo RJ, Plautz EJ, Frost SB. (2001) Role of adaptive plasticity in recovery of function after damage to motor cortex. Muscle Nerve 24(8): 1000-19. |
Figures(1)

Wanda M. Snow, Brenda M. Stoesz, Judy E. Anderson. The Cerebellum in Emotional Processing: Evidence from Human and Non-Human Animals[J]. AIMS Neuroscience, 2014, 1(1): 96-119. doi: 10.3934/Neuroscience.2014.1.96







 DownLoad:
DownLoad: