Citation: Daniel Amsterdam. Unique natural and adaptive response mechanisms to control and eradicate HIV infection[J]. AIMS Allergy and Immunology, 2018, 2(3): 113-125. doi: 10.3934/Allergy.2018.3.113

| [1] | Center for Disease Control and Prevention (1981) Pneumocystis pneumonia. MMWR 30: 1–3. |
| [2] | Center for Disease Control and Prevention (2015) Prevalence of diagnosed and undiagnosed HIV infection-2008–2012. MMWR 64: 657–662. |
| [3] | Wainberg MA, Zaharatos GJ, Brenner BG (2010) Development of antiretroviral drug resistance. N Eng J Med 365: 637–645. |
| [4] |
Maartens G, Celum C, Lewin SR (2014) HIV infection: epidemiology, pathogenesis, treatment, and prevention. Lancet 384: 258–271. doi: 10.1016/S0140-6736(14)60164-1

|
| [5] |
Huang Z, Tomitaka A, Raymond A, et al. (2017) Current application of CRISPR/Cas9 gene-editing technique to eradication of HIV/AIDS. Gene Ther 24: 377–384. doi: 10.1038/gt.2017.35
|
| [6] |
Chun TW, Engel D, Mizell SB, et al. (1998) Induction of HIV-1 replication in latently infected CD4+ T cells using a combination of cytokines. J Exp Med 188: 83–91. doi: 10.1084/jem.188.1.83
|
| [7] |
Finzi D, Hemankova M, Pierson T, et al. (1997) Identification of a reservoir for HIV-1 in patients on highly active antiretroviral therapy. Science 278: 1295–1300. doi: 10.1126/science.278.5341.1295
|
| [8] |
McElrath MJ, Steinman RM, Cohn ZA, et al. (1991) Latent HIV-1 infection in enriched populations of blood monocytes and T cells from seropositive patients. J Clin Inv 87: 27–30. doi: 10.1172/JCI114981
|
| [9] |
Chun TW, Engel D, Berrey MM, et al. (1998) Early establishment of a pool of latently infected, resting CD4 (+) T cells during primary HIV infection. PNAS 95: 8869–8873. doi: 10.1073/pnas.95.15.8869
|
| [10] |
Bagasra O, Lavi E, Bobroski I, et al. (1996) Cellular reservoirs of HIV-1 in the central nervous system of infected individuals: identification by the combination of in situ polymerase chain reaction and immunohistochemistry. AIDS 10: 573–585. doi: 10.1097/00002030-199606000-00002
|
| [11] |
Fischer-Smith T, Croul S, Sverstiuk AE, et al. (2001) CNS invasion by CD14+/CD16+ peripheral blood derived monocytes in HIV dementia: perivascular accumulation and reservoir of HIV infection. J Neurovirol 7: 528–541. doi: 10.1080/135502801753248114
|
| [12] |
Petito CK, Chen H, Mastri AR, et al. (1999) HIV infection of choroid plexus in AIDS and asymptomatic HIV infected patients suggests that choroid plexus might be a good reservoir of productive infection. J Neurovirol 5: 670–677. doi: 10.3109/13550289909021295
|
| [13] |
Chun TW, Carruth L, Finzi D, et al. (1997) Quantification of latent tissue reservoirs and total boy viral load in HIV-1 infection. Nature 387: 183–188. doi: 10.1038/387183a0
|
| [14] |
Chun TW, Nickle DC, Justement JS, et al. (2008) Persistence of HIV in gut-associated lymphoid tissue despite long-term antiretroviral therapy. J Inf Dis 197: 714–720. doi: 10.1086/527324
|
| [15] |
Smith PD, Meng G, Salazar-Gonzalez JF, et al. (2003) Macrophage HIV-1 infection and the potential gastrointestinal tract reservoir. J Leuk Bio 74: 642–649. doi: 10.1189/jlb.0503219
|
| [16] |
Lambert-Niclot S, Peytavin G, Duvivier C, et al. (2010) Low frequency of intermittent HIV-1 semen excretion on patients treated with darunavir-ritonavir at 600/100 milligrams twice a day plus two nucleoside reverse transcriptase inhibitors or monotherapy. Antimicrob Agents Chemo 54: 4910–4913. doi: 10.1128/AAC.00725-10
|
| [17] |
Cu-Uvin S, DeLong AK, Venkatesh KK, et al. (2010) Genital track HIV-1 RNA shedding among women with below detectable plasma viral load. AIDS 24: 2489–2497. doi: 10.1097/QAD.0b013e32833e5043
|
| [18] |
Hutter G, Nowak D, Mossner M, et al. (2009) Long term control of HIV by CCR5 Delta 32/Delta 32 stem-cell transplantation. N Engl J Med 360: 692–968. doi: 10.1056/NEJMoa0802905
|
| [19] |
Perelson AS, Esunger P, Cao Y, et al. (1997) Decay characteristics of HIV-1 infected compartments during combination therapy. Nature 387: 188–191. doi: 10.1038/387188a0
|
| [20] | Evering TH, Mehandru S, Racz P, et al. (2012) Absence of HIV-1 evolution in the gut-associated lymphoid tissue from patients on combination antiviral therapy initiated during primary infection. PLoS Pathog 8: e1002306. |
| [21] |
Kearney MF, Spindler J, Shao W, et al. (2014) Lack of detectable HIV-1 molecular evolution during suppressive antiretroviral therapy. PLoS Pathog 10: e1004010. doi: 10.1371/journal.ppat.1004010
|
| [22] | Cáceres CF, Mayer KH, Baggaley R, et al. (2015) PrEP implementation science: state-of-the-art and research agenda. J Int Aids Soc 18: 20527. |
| [23] | World Health Organization (2017) Implementation tool for pre-exposure prophylaxis (PrEP) of HIV infection. Policy Brief WHO Reference number: WHO/HIV/2017.19. |
| [24] |
Chomont N, El-Far M, Ancuta P, et al. (2009) HIV reservoir size and persistence are driven by T cell survival and homeostatic proliferation. Nat Med 15: 893–900. doi: 10.1038/nm.1972
|
| [25] |
Kulpa DA, Lawani M, Cooper A, et al. (2013) PD-1 co-inhibitory signals: the link between pathogenesis and protection. Semin Immunol 25: 219–227. doi: 10.1016/j.smim.2013.02.002
|
| [26] |
Porichis F, Kaufmann DE (2012) Role of PD-1 in HIV-1 pathogenesis and as a target for therapy. Curr HIV-AIDS Rep 9: 81–90. doi: 10.1007/s11904-011-0106-4
|
| [27] |
Reguzova AY, Karpenko LI, Mechetina LV, et al. (2015) Peptide MHC multimer-based monitoring of CD8 T cells in HIV-1 infection and HIV vaccine development. Expert Rev Vaccines 14: 69–84. doi: 10.1586/14760584.2015.962520
|
| [28] |
Zhang JY, Zhang Z, Wang XZ, et al. (2007) PD-1 upregulation is correlated with HIV specific memory CD8 (+) T cells exhaustion in typical progressors but not in long-term non-progressors. Blood 109: 4671–4678. doi: 10.1182/blood-2006-09-044826
|
| [29] |
Cohen MS, Chen YQ, McCauley M, et al. (2016) Antiretroviral therapy for the prevention of HIV-1 transmission. N Engl J Med 375: 830–839. doi: 10.1056/NEJMoa1600693
|
| [30] |
Chun TW, Stuyver L, Misell SB, et al. (1997) Presence of an inducible HIV-1 latent reservoir during highly active antiretroviral therapy. PNAS 94: 13193–13197. doi: 10.1073/pnas.94.24.13193
|
| [31] |
Wong JK, Hezareh M, Gunthard HF, et al. (1997) Recovery of replication-competent HIV despite prolonged suppression of plasma viremia. Science 278: 1291–1295. doi: 10.1126/science.278.5341.1291
|
| [32] |
Finzi D, Blankson I, Siliciano JD, et al. (1999) Latent infection of CD4+ T cells provides a mechanism for lifelong persistence of HIV-1, even in patients on effective combination therapy. Nat Med 5: 512–517. doi: 10.1038/8394
|
| [33] |
Deeks SG (2012) HIV: shock and kill. Nature 487: 439–440. doi: 10.1038/487439a
|
| [34] |
Siliciano JD, Kajdas J, Finzi D, et al. (2003) Long-term follow-up studies confirm the stability of the latent reservoir for HIV-1 in resting CD4+ T cells. Nat Med 9: 727–728. doi: 10.1038/nm880
|
| [35] |
Ruelas DS, Greene WC (2013) An integrated overview of HIV-1 latency. Cell 155: 519–529. doi: 10.1016/j.cell.2013.09.044
|
| [36] |
Donahue DA, Wainberg MA (2013) Cellular and molecular mechanisms involved in the establishment of HIV-1 latency. Retrovirology 10: 11. doi: 10.1186/1742-4690-10-11
|
| [37] |
Strain MC, Little SJ, Daar ES, et al. (2005) Effect of treatment during primary infection on establishment and clearance of cellular reservoirs of HIV-1. J Infect Dis 191: 1410–1418. doi: 10.1086/428777
|
| [38] | Procopio FA, Fromentin R, Kulpa DA, et al. (2015) A novel assay to measure the magnitude of the inducible viral reservoir in HIV infected individuals. Ebiomedicine 2: 872–881. |
| [39] | Walker B, McMichael A (2012) The T-cell response to HIV. Cold Spring Harb Perspect Med 2: e007054. |
| [40] | Lia MK, Hawkins N, Ritchie AI, et al. (2013) CHAVI Core B. Vertical T cell immunodominance and epitope entropy determine HIV-1 escape. J Clin Invest 123: 380–393. |
| [41] |
Ferrari G, Korber B, Goonetilde N, et al. (2011) Relationship between functional profile of HIV-1 specific CD8 T cells and epitope variability with the selection of escape mutants in acute HIV-1 infection. PLoS Pathog 7: e1001273. doi: 10.1371/journal.ppat.1001273
|
| [42] |
Streeck H, Brumme ZL, Anastario M, et al. (2008) Antigen load and viral sequence diversification determine the functional profile of HIV-1 specific CD8+ T cells. PLoS Med 5: e100. doi: 10.1371/journal.pmed.0050100
|
| [43] |
Pollana J, Nonsignori M, Moody MA, et al. (2013) Epitope specificity of human immunodeficiency virus-1 antibody dependent cellular cytotoxicity (ADCC) responses. Curr HIV Res 11: 378–387. doi: 10.2174/1570162X113116660059
|
| [44] |
Liao HX, Bonsignori M, Alam SM, et al. (2013) Vaccine induction of antibodies against a structurally heterogeneous site of immune pressure within HIV-1 envelope protein variable regions 1 and 2. Immunity 38: 176–186. doi: 10.1016/j.immuni.2012.11.011
|
| [45] |
Mascola JR, D'Souza P, Gilbert, et al. (2005) Recommendations for the design and use of standard virus panels to assess neutralizing antibody responses elicited by candidate human immunodeficiency virus type 1 vaccines. J Virol 79: 10103–10107. doi: 10.1128/JVI.79.16.10103-10107.2005
|
| [46] |
Xu L, Pegu A, Rao E, et al. (2017) Trispecific broadly neutralizing HIV antibodies mediate potent SHIV protection in macaques. Science 358: 85–90. doi: 10.1126/science.aan8630
|
| [47] |
Julg B, Liu PT, Wagh K, et al. (2017) Protection against a mixed SHIV challenge by a broadly neutralizing antibody cocktail. Sci Trans Med 9: eaao4235. doi: 10.1126/scitranslmed.aao4235
|
| [48] |
McCoy LE, Burton DR (2017) Identification and specificity of broadly neutralizing antibodies against HIV. Immunol Rev 275: 11–20. doi: 10.1111/imr.12484
|
| [49] |
Amsterdam D (2015) Immunotherapeutic approaches for the control and eradication of HIV. Immunol Inv 44: 719–730. doi: 10.3109/08820139.2015.1096680
|
| [50] |
Hirsch AJ (2010) The use of RNAi-based screens to identify host proteins involved in viral replication. Future Microbiol 5: 303–311. doi: 10.2217/fmb.09.121
|
| [51] |
Jacqua JM, Triques K, Stevenson M (2002) Modulation of HIV-1 replication by RNA interference. Nature 418: 435–438. doi: 10.1038/nature00896
|
| [52] |
Nishitsuji H, Kohara M, Kannagi M, et al. (2006) Effective suppression of human immunodeficiency virus type 1 through a combination of short- and long-hairpin RNAs targeting essential sequences for retroviral integration. J Virol 80: 7658–7666. doi: 10.1128/JVI.00078-06
|
| [53] |
Suzuki K, Ishida T, Yamagishi M, et al. (2011) Transcriptional gene silencing of HIV-1 through promoter targeting RNA is highly specific. RNA Biol 8: 1035–1046. doi: 10.4161/rna.8.6.16264
|
| [54] | Taksuchi Y, Nagumo T, Hashino H (1988) Low fidelity of cell-free DNA synthesis by reverse transcriptase of human immunodeficiency virus. J Virol 62: 3000–3002. |
| [55] |
Knoepfel SA, Centlivre M, Liu YP, et al. (2012) Selection of RNAi-based inhibitors for anti-HIV gene therapy. World J Virol 1: 79–90. doi: 10.5501/wjv.v1.i3.79
|
| [56] |
Anderson J, Akkina R (2005) CXCR4 and CCR5 shRNA transgenic CD34+ cell derived macrophages are functionally normal resist HIV-1 infection. Retrovirology 2: 53. doi: 10.1186/1742-4690-2-53
|
| [57] | Martinez MA, Clotet B, Este JA (2002) RNA interference of HIV replication. Trends Immunol 3: 559–561. |
| [58] |
Boutimah F, Eekels JJ, Liu YP, et al. (2013) Antiviral strategy combining antiretroviral drugs with RNAi-mediated attack on HIV-1 and cellular co-factors. Antiviral Res 98: 121–129. doi: 10.1016/j.antiviral.2013.02.011
|
| [59] |
Wolstein O, Boyd M, Millington M, et al. (2014) Preclinical safety and efficacy of an anti-HIV-1 lentiviral vector containing a short hairpin RNA to CCR5 and the C46 fusion inhibitor. Mol Ther Meth Clin Dev 1: 11. doi: 10.1038/mtm.2013.11
|
| [60] |
Cox DB, Platt RJ, Zhang F (2015) Therapeutic genome editing: Prospects and challenges. Nat Med 21: 121–131. doi: 10.1038/nm.3793
|
| [61] |
Badia R, Rivera-Munoz E, Clotet B, et al. (2014) Gene editing using a zinc-finger nuclease mimicking the CCR5Delta32 mutation induces resistance to CCR5-using HIV-1. J Antimicrob Chemother 69: 1755–1759. doi: 10.1093/jac/dku072
|
| [62] |
Perez EE, Wang J, Miller JC, et al. (2008) Establishment of HIV-1 resistance in CD4+ T cells by genome editing using zinc-finger nucleases. Nat Biotechnol 26: 808–816. doi: 10.1038/nbt1410
|
| [63] |
Yao Y, Nashun B, Zhou T, et al. (2012) Generation of CD34+ cells from CCR5-disrupted human embryonic and induced pluripotent stem cells. Hum Gene Ther 23: 238–242. doi: 10.1089/hum.2011.126
|
| [64] |
Wilen CB, Wang J, Tilton JC, et al. (2011) Engineering HIV-resistant human CD4+ T cells with CXCR4-specific zinc-finger nucleases. PLoS Pathog 7: e1002020. doi: 10.1371/journal.ppat.1002020
|
| [65] |
Didigu CA, Wilen CB, Wang J, et al. (2014) Simultaneous zinc-finger nuclease editing of HIV coreceptors ccr5 and cxcr4 protects CD4+ T cells from HIV-1 infection. Blood 123: 61–69. doi: 10.1182/blood-2013-08-521229
|
| [66] | Shi B, Li J, Shi X, et al. (2017) TALEN-mediated knockout of CCR5 confers protection against infection of human immunodeficiency virus. J AIDS 74. |
| [67] |
Tebas P, Stein D, Tang WW, et al. (2014) Gene editing of CCR5 in autologous CD4 T cells of persons infected with HIV. N Engl J Med 370: 901–910. doi: 10.1056/NEJMoa1300662
|
| [68] |
Mealer DA, Brennan AL, Jiang S, et al. (2013) Efficient clinical scale gene modification via zinc finger nuclease-targeted disruption of the HIV co-receptor CCR5. Hum Gene Ther 24: 245–258. doi: 10.1089/hum.2012.172
|
| [69] |
Strong CL, Guerra HP, Mathew KR, et al. (2015) Damaging the integrated HIV proviral DNA with TALENs. PLoS One 10: e0125652. doi: 10.1371/journal.pone.0125652
|
| [70] |
Horvath P, Barrangou R (2010) CRISPR/Cas, the immune system of bacteria and archaea. Science 327: 167–170. doi: 10.1126/science.1179555
|
| [71] |
Cong L, Ran FA, Cox D, et al. (2013) Multiplex genome engineering using CRISP/Cas systems. Science 339: 819–823. doi: 10.1126/science.1231143
|
| [72] |
Hsu PD, Lander ES, Zhang F (2014) Development and applications of CRISPR-Case9 for genome engineering. Cell 157: 1262–1278. doi: 10.1016/j.cell.2014.05.010
|
| [73] |
Ebina H, Misawa N, Kanemura Y, et al. (2013) Harnessing the CRIPSR/Cas 9 system to disrupt latent HIV-1 provirus. Sci Rep 3: 2510. doi: 10.1038/srep02510
|
| [74] |
Hou P, Chen S, Wang S, et al. (2015) Genome editing of CXCR4 by CRISPR/Cas9 confers cells resistant to HIV-1 infection. Sci Rep 5: 15577. doi: 10.1038/srep15577
|
| [75] |
Ye L, Wang J, Beyer AI, et al. (2014) Seamless modification of wild-type induced pluripotent stem cells to the natural CCR5Delta32 mutation confers resistance to HIV infection. PNAS 111: 9591–9596. doi: 10.1073/pnas.1407473111
|
| [76] | Dampier W, Nonnemacher MR, Sullivan NT, et al. (2014) HIV excision utilizing CRISPR/Cas 9 technology: attacking the proviral quasispecies in reservoirs to achieve a cure. MOJ Immunol 1: 00022. |
| [77] |
Zhu W, Lei R, De Duff Y, et al. (2015) The CRISPR/Cas9 system inactivates latent HIV-1 proviral DNA. Retrovirology 12: 22. doi: 10.1186/s12977-015-0150-z
|
| [78] |
Wang Z, Pan Q, Gendron P, et al. (2016) CRISPR/Cas-9-derived mutations both inhibit HIV-1 replication and accelerate viral escape. Cell Rep 15: 481–489. doi: 10.1016/j.celrep.2016.03.042
|
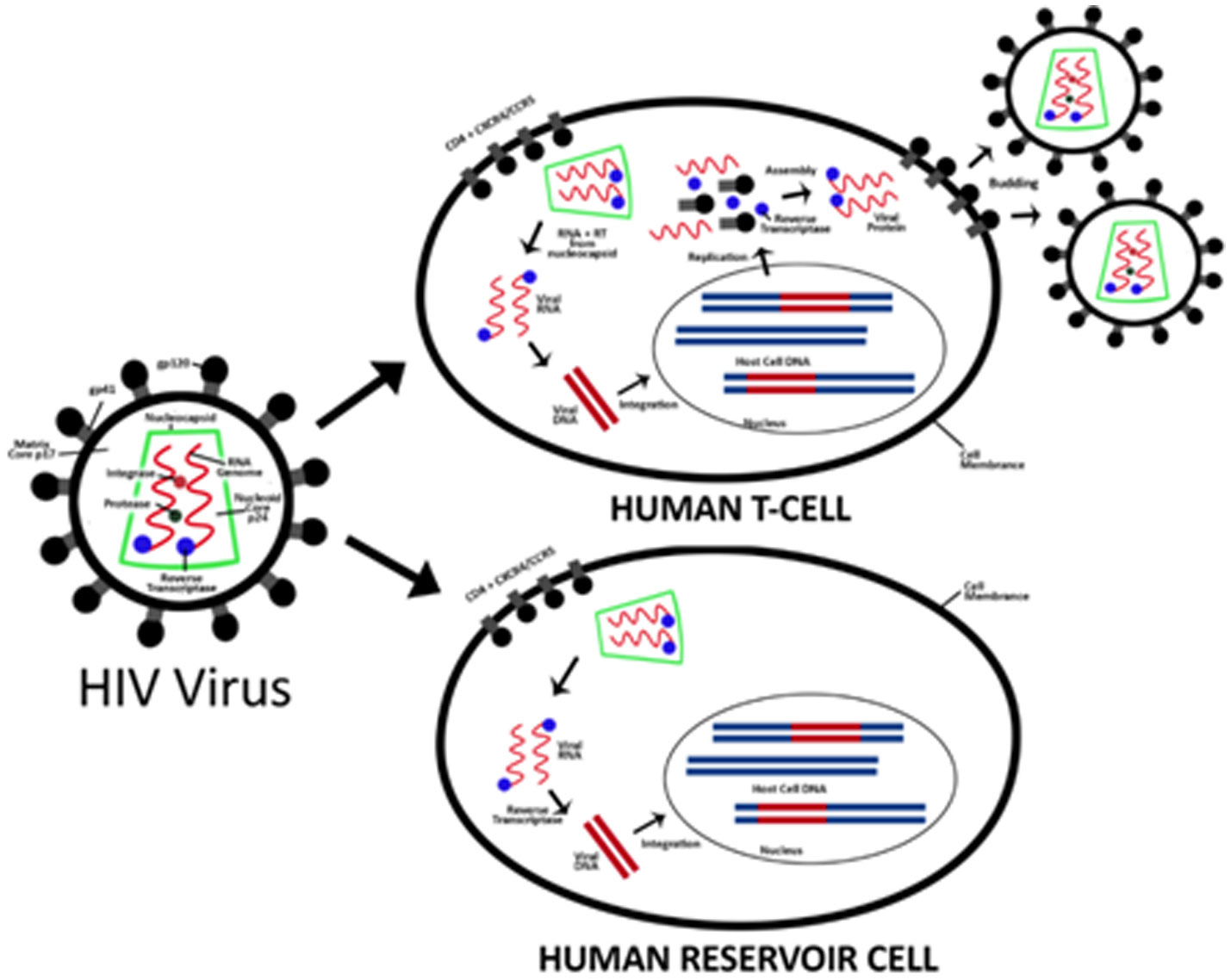
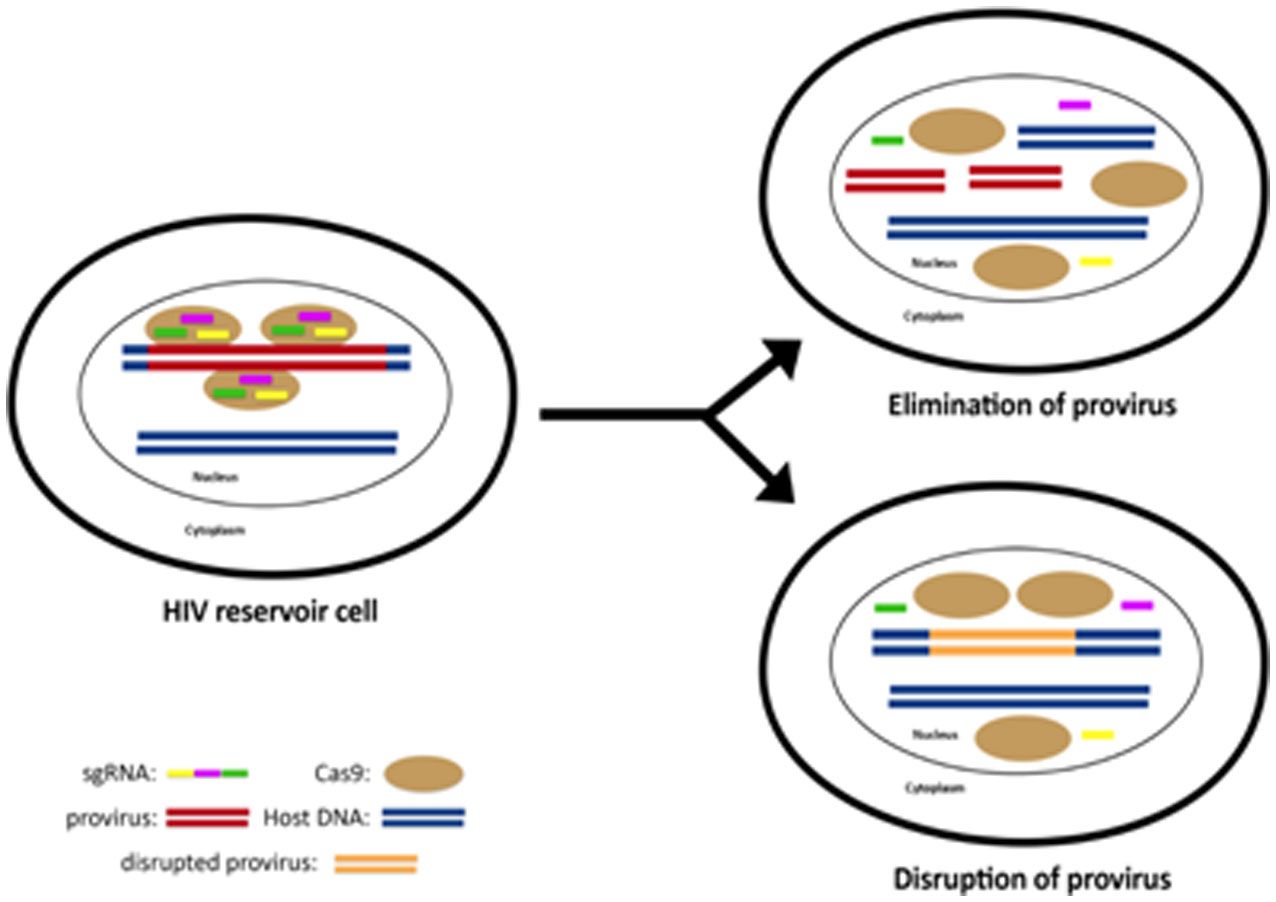
Figures(2)

Daniel Amsterdam. Unique natural and adaptive response mechanisms to control and eradicate HIV infection[J]. AIMS Allergy and Immunology, 2018, 2(3): 113-125. doi: 10.3934/Allergy.2018.3.113







 DownLoad:
DownLoad: