Citation: Nikolina-Alexandra Xaxiri, Eleni Nikouli, Panagiotis Berillis, Konstantinos Ar. Kormas. Bacterial biofilm development during experimental degradation of Melicertus kerathurus exoskeleton in seawater[J]. AIMS Microbiology, 2018, 4(3): 397-412. doi: 10.3934/microbiol.2018.3.397

| [1] | Zobell CE, Rittenberg SC (1938) The occurrence and characteristics of chitinoclastic bacteria in the sea. J Bacteriol 35: 275–287. |
| [2] |
Gooday GW (1990) The ecology of chitin degradation. Adv Microb Ecol 11: 387–430. doi: 10.1007/978-1-4684-7612-5_10

|
| [3] | Alluwihare LI, Meador T (2008) Chemical composition of marine dissolved organic nitrogen, In: Capone DG, Bronk DA, Mulholland MR, Carpenter EJ, Editors, Nitrogen in the marine environment, Academic Press, 95–140. |
| [4] |
Souza CP, Almeida BC, Colwell RR, et al. (2011) The importance of chitin in the marine environment. Mar Biotechnol 13: 823. doi: 10.1007/s10126-011-9388-1
|
| [5] |
Seki H (1965) Rough estimation on chitin decomposition in the ocean. J Oceanogr Soc Jpn 21: 253–260. doi: 10.5928/kaiyou1942.21.253
|
| [6] |
Gooday GW (1990) Physiology of microbial degradation of chitin and chitosan. Biodegradation 1: 177–190. doi: 10.1007/BF00058835
|
| [7] |
Teng WL, Khor E, Tan TK, et al. (2001) Concurrent production of chitin from shrimp shells and fungi. Carbohyd Res 332: 305–316. doi: 10.1016/S0008-6215(01)00084-2
|
| [8] | FAO (2017) An overview of recently published global aquaculture statistics. FAO aquaculture newsletter No. 56, 2017. |
| [9] |
Zakaria Z, Hall GM, Shama G (1998) Lactic acid fermentation of scampi waste in a rotating horizontal bioreactor for chitin recovery. Process Biochem 33: 1–6. doi: 10.1016/S0032-9592(97)00069-1
|
| [10] |
No HK, Meyers SP (1995) Preparation and characterization of chitin and chitosan-a review. J Aquat Food Prod T 4: 27–52. doi: 10.1300/J030v04n02_03
|
| [11] |
Kurita K (2001) Controlled functionalization of the polysaccharide chitin. Prog Polym Sci 26: 1921–1971. doi: 10.1016/S0079-6700(01)00007-7
|
| [12] |
Healy M, Green A, Healy A (2003) Bioprocessing of marine crustacean shell waste. Acta Biotechnol 23: 151–160. doi: 10.1002/abio.200390023
|
| [13] |
Islam MS, Khan S, Tanaka M (2004) Waste loading in shrimp and fish processing effluents: Potential source of hazards to the coastal and nearshore environments. Mar Pollut Bull 49: 103–110. doi: 10.1016/j.marpolbul.2004.01.018
|
| [14] | Beaney P, Lizardi-Mendoza J, Healy M (2010) Comparison of chitins produced by chemical and bioprocessing methods. J Chem Technol Biotechnol 80: 145–150. |
| [15] |
Brzezinska MS, Jankiewicz U, Burkowska A, et al. (2014) Chitinolytic microorganisms and their possible application in environmental protection. Curr Microbiol 68: 71–81. doi: 10.1007/s00284-013-0440-4
|
| [16] | Brück WM, Slater JW, Carney BF (2010) Chitin and chitosan from marine organisms, In: Kim SK, Editor, Chitin, chitosan, oligosaccharides and their derivatives, Biological activities and applications, Boca Raton: CRC Press, 11–24. |
| [17] | Jo GH, Park RD, Jung WJ (2011) Enzymatic production of chitin from crustacean shell waste, In: Kim SK, Editor, Chitin, chitosan, oligosaccharides and their derivatives, biological activities and applications, Boca Raton, CRC Press, 37–46. |
| [18] |
Sogin ML, Morrison HG, Huber JA, et al. (2006) Microbial diversity in the deep sea and the underexplored "rare biosphere". P Natl Acad Sci USA 103: 12115–12120. doi: 10.1073/pnas.0605127103
|
| [19] | FAO, 2018. Available from: http://www.fao.org/fishery/species/2587/en. |
| [20] |
Klindworth A, Pruesse E, Schweer T, et al. (2012) Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res 41: e1. doi: 10.1093/nar/gks1297
|
| [21] |
Schloss PD, Westcott SL, Ryabin T, et al. (2009) Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Microb 75: 7537–7541. doi: 10.1128/AEM.01541-09
|
| [22] |
Quince C, Lanzen A, Curtis TP, et al. (2009) Accurate determination of microbial diversity from 454 pyrosequencing data. Nat Methods 6: 639–641. doi: 10.1038/nmeth.1361
|
| [23] |
Pruesse E, Quast C, Knittel K, et al. (2007) SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res 35: 7188–7196. doi: 10.1093/nar/gkm864
|
| [24] |
Kunin V, Engelbrektson A, Ochman H, et al. (2010) Wrinkles in the rare biosphere: Pyrosequencing errors can lead to artificial inflation of diversity estimates. Environ Microbiol 12: 118–123. doi: 10.1111/j.1462-2920.2009.02051.x
|
| [25] |
Stackenbrandt E, Goebel BM (1994) Taxonomic note: A place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int J Syst Evol Micr 44: 846–849. doi: 10.1099/00207713-44-4-846
|
| [26] |
Newton RJ, Shade A (2016) Lifestyles of rarity: Understanding heterotrophic strategies to inform the ecology of the microbial rare biosphere. Aquat Microb Ecol 78: 51–63. doi: 10.3354/ame01801
|
| [27] | Brenner DJ, Krieg NR, Staley JT (2010a) Bergey's manual of systematic bacteriology. Part B: The Gammaproteobacteria, New York: Springer, 1106. |
| [28] | Brenner DJ, Krieg NR, Staley JT (2010b) Bergey's manual of systematic bacteriology. Part C: The Alpha-, Beta-, Delta-, and Epsilonproteobacteria, New York: Springer, 1388. |
| [29] | Krieg NR, Staley JT, Brown DR, et al. (2010) Bergey's manual of systematic bacteriology. The Bacteroidetes, Spirochaetes, Tenericutes (Mollicutes), Acidobacteria, Fibrobacteres, Fusobacteria, Dictyoglomi, Gemmatimonadetes, Lentisphaerae, Verrucomicrobia, Chlamydiae, and Planctomycetes, New York: Springer, 949. |
| [30] |
Roller BRK, Stoddard SF, Schmidt TM (2016) Exploiting rRNA operon copy number to investigate bacterial reproductive strategies. Nat Microbiol 1: 16160. doi: 10.1038/nmicrobiol.2016.160
|
| [31] | Yan YY, Zhu FF, Sun QQ, Pseudoalteromonas porphyrae sp. nov., a marine bacterium that is the causative agent of bud blight disease of Porphyrae yezoensis, 2002. Available from: https://www.ncbi.nlm.nih.gov. |
| [32] |
Dimitrieva GY, Crawford RL, Yüksel GÜ (2006) The nature of plant growth-promoting effects of a pseudoalteromonad associated with the marine algae Laminaria japonica and linked to catalase excretion. J Appl Microbiol 100: 1159–1169. doi: 10.1111/j.1365-2672.2006.02831.x
|
| [33] | Kacar A, Kocyigit A, Ozdemir G, et al. (2009) The development of biofilm bacteria on panels coated by different antifouling paints in the marinas. Fresen Environ Bull 18: 2004–2012. |
| [34] | Lee RD, Jospin G, Lang JM, et al. (2016) Draft genome sequences of two Pseudoalteromonas porphyrae strains isolated from seagrass sediment. Genome Announc 4: e00092–e00116. |
| [35] |
Offret C, Desriac F, Le Chevalier P, et al. (2016) Spotlight on antimicrobial metabolites from the marine bacteria Pseudoalteromonas: Chemodiversity and ecological significance. Mar Drugs 14: 129. doi: 10.3390/md14070129
|
| [36] | Valdenegro-Vega V, Naeem S, Carson J, et al. (2013) Culturable microbiota of ranched southern bluefin tuna (Thunnus maccoyii Castelnau). J Appl Microbiol 115: 923–932. |
| [37] |
Li J, Tan B, Mai K (2011) Isolation and identification of a bacterium from marine shrimp digestive tract: A new degrader of starch and protein. J Ocean U China 10: 287–292. doi: 10.1007/s11802-011-1849-7
|
| [38] | Ivanova EP, Sawabe T, Lysenko AM, et al. (2002) Pseudoalteromonas ruthenica sp. nov., isolated from marine invertebrates. Int J Syst Evol Microbiol 52: 235–240. |
| [39] |
Vynne NG, Mansson M, Nielsen KF, et al. (2011) Bioactivity, chemical profiling, and 16S rRNA-based phylogeny of Pseudoalteromonas strains collected on a global research cruise. Mar Biotechnol 13: 1062–1073. doi: 10.1007/s10126-011-9369-4
|
| [40] | Ahmad A, Sulieman MFI, Usup G (2016) Anti-biofilm activity of the marine bacterium Pseudoalteromonas ruthenica KLPp3 against Serratia marcescens and Vibrio alginolyticus. Malays J Microbiol 12: 30–34. |
| [41] |
Sass AM, Sass H, Coolen MJL, et al. (2001) Microbial communities in the chemocline of a hypersaline deep-sea basin (Urania Basin, Mediterranean Sea). Appl Environ Microb 67: 5392–5402. doi: 10.1128/AEM.67.12.5392-5402.2001
|
| [42] |
Ettoumi B, Bouhajja E, Borin S, et al. (2010) Gammaproteobacteria occurrence and microdiversity in Tyrrhenian Sea sediments as revealed by cultivation-dependent and -independent approaches. Syst Appl Microbiol 33: 222–231. doi: 10.1016/j.syapm.2010.02.005
|
| [43] |
De Vitis V, Guidi B, Contente ML, et al. (2015) Marine microorganisms as source of stereoselective esterases and ketoreductases: Kinetic resolution of a prostaglandin intermediate. Mar Biotechnol 17: 144–152. doi: 10.1007/s10126-014-9602-z
|
| [44] |
Qurashi AW, Sabri AN (2012) Biofilm formation in moderately halophilic bacteria is influenced by varying salinity levels. J Basic Microbiol 52: 566–572. doi: 10.1002/jobm.201100253
|
| [45] |
Kasan NA, Ghazali NA, Ikhwanuddin M, et al. (2017) Isolation of potential bacteria as inoculum for biofloc formation in pacific whiteleg shrimp, litopenaeus vannamei culture ponds. Pak J Biol Sci 20: 306–313. doi: 10.3923/pjbs.2017.306.313
|
| [46] | Suantika G, Aditiawati P, Astuti DI, et al. (2013) The use of indigenous probiotic halomonas aquamarina and shewanella algae for white shrimp (Litopenaeus vannamei boone) hatchery productivity in zero water discharge system. J Aquac Res Dev 4: 194. |
| [47] |
Inbakandan D, Murthy PS, Venkatesan R, et al. (2010) 16S rDNA sequence analysis of culturable marine biofilm forming bacteria from a ship's hull. Biofouling 26: 893–899. doi: 10.1080/08927014.2010.530347
|
| [48] |
Sorkhoh NA, Alawadhi H, Almailem DM, et al. (2010) Agarolytic bacteria with hydrocarbon-utilization potential in fouling material from the Arabian Gulf coast. Int Biodeter Biodegr 64: 554–559. doi: 10.1016/j.ibiod.2010.06.007
|
| [49] |
Al-Mailem DM, Eliyas M, Radwan SS (2012) Enhanced haloarchaeal oil removal in hypersaline environments via organic nitrogen fertilization and illumination. Extremophiles 16: 751–758. doi: 10.1007/s00792-012-0471-y
|
| [50] | Choi A, Cho JC (2010) Reinekea aestuarii sp. nov., isolated from tidal flat sediment. Int J Syst Evol Microbiol 60: 2813–2817. |
| [51] |
Avci B, Hahnke RL, Chafee M, et al. (2017) Genomic and physiological analyses of "Reinekea forsetii" reveal a versatile opportunistic lifestyle during spring algae blooms. Environ Microbiol 19: 1209–1221. doi: 10.1111/1462-2920.13646
|
| [52] | Choi EJ, Kwon HC, Koh HY, et al. (2010) Colwellia asteriadis sp. nov., a marine bacterium isolated from the starfish Asterias amurensis. Int J Syst Evol Microbiol 60: 1952–1957. |
| [53] | Gillan DC, Speksnijder AG, Zwart G, et al. (1998) Genetic diversity of the biofilm covering Montacuta ferruginosa (Mollusca, bivalvia) as evaluated by denaturing gradient gel electrophoresis analysis and cloning of PCR-amplified gene fragments coding for 16S rRNA. Appl Environ Microb 64: 3464–3472. |
| [54] |
Schwermer CU, Lavik G, Abed RM, et al. (2008) Impact of nitrate on the structure and function of bacterial biofilm communities in pipelines used for injection of seawater into oil fields. Appl Environ Microb 74: 2841–2851. doi: 10.1128/AEM.02027-07
|
| [55] | Finnegan L, Garciamelgares M, Gmerek T, et al. (2011) A survey of culturable aerobic and anaerobic marine bacteria in de novo biofilm formation on natural substrates in St. Andrews Bay, Scotland. Anton Leeuw 100: 399–404. |
| [56] |
Casillo A, Lanzetta R, Parrilli M, et al. (2018) Exopolysaccharides from marine and marine extremophilic bacteria: Structures, properties, ecological roles and applications. Mar Drugs 16: 69. doi: 10.3390/md16020069
|
| [57] | Kim SJ, Tamura T, Hamada M, et al. (2012) Compostimonas suwonensis gen. nov., sp. nov., isolated from spent mushroom compost. Int J Syst Evol Microbiol 62: 2410–2416. |
| [58] |
Tiwari K, Gupta RK (2013) Diversity and isolation of rare actinomycetes: An overview. Crit Rev Microbiol 39: 256–294. doi: 10.3109/1040841X.2012.709819
|
| [59] | Bernardet JF, Bowman JP (2006) The Genus Flavobacterium, In: Dworkin M, Falkow S, Rosenberg E, Schleifer KH, Stackebrandt E, Editors, The Prokaryotes, New York: Springer, 2006. |
| [60] |
Doghri I, Rodrigues S, Bazire A, et al. (2015) Marine bacteria from the French Atlantic coast displaying high forming-biofilm abilities and different biofilm 3D architectures. BMC Microbiol 15: 231. doi: 10.1186/s12866-015-0568-4
|
| [61] |
Martins P, Cleary DFR, Pires ACC, et al. (2013) Molecular analysis of bacterial communities and detection of potential pathogens in a recirculating aquaculture system for Scophthalmus maximus and Solea senegalensis. PLoS One 8: e80847. doi: 10.1371/journal.pone.0080847
|
| [62] |
Fernández-Álvarez C, Torres-Corral Y, Saltos-Rosero N, et al. (2017) MALDI-TOF mass spectrometry for rapid differentiation of Tenacibaculum species pathogenic for fish. Appl Microbiol Biotechnol 101: 5377–5390. doi: 10.1007/s00253-017-8324-3
|
| [63] | Banning EC, Casciotti KL, Kujawinski EB (2010) Novel strains isolated from a coastal aquifer suggest a predatory role for flavobacteria. FEMS Microbiol Ecol 73: 254–270. |
| [64] | Faury N, Saulnier D, Thompson FL, et al. (2004) Vibrio crassostreae sp. nov., isolated from the haemolymph of oysters (Crassostrea gigas). Int J Syst Evol Microbiol 54: 2137–2140. |
| [65] | Poli A, Romano I, Mastascusa V, et al. (2018) Vibrio coralliirubri sp. nov., a new species isolated from mucus of red coral (Corallium rubrum) collected at Procida island, Italy. Anton Leeuw 2018: 1–11. |
| [66] |
Rizzo L, Fraschetti S, Alifano P, et al. (2016) Association of Vibrio community with the Atlantic Mediterranean invasive alga Caulerpa cylindracea. J Exp Mar Biol Ecol 475: 129–136. doi: 10.1016/j.jembe.2015.11.013
|
| [67] | Romalde JL, Dieguez AL, Lasa A, et al. (2013) New Vibrio species associated to molluscan microbiota: A review. Front Microbiol 4: 413. |
| [68] |
Stabili L, Giangrande A, Pizzolante G, et al. (2014) Characterization of vibrios diversity in the mucus of the polychaete Myxicola infundibulum (Annellida, Polichaeta). Microb Ecol 67: 186–194. doi: 10.1007/s00248-013-0312-2
|
| [69] |
Bruto M, James A, Petton B, et al. (2017) Vibrio crassostreae, a benign oyster colonizer turned into a pathogen after plasmid acquisition. ISME J 11: 1043–1052. doi: 10.1038/ismej.2016.162
|
| [70] | Huang D, Liang X, Peng L, et al. (2017) Effects of Vibrio biofilms of different sources on settlement of plantigrades of the mussel Mytilus coruscus. J Fish China 41: 1149–1147. |
| [71] |
Sugita H, Mizuki H, Itoi S (2012) Diversity of siderophore-producing bacteria isolated from the intestinal tracts of fish along the Japanese coast. Aquac Res 43: 481–488. doi: 10.1111/j.1365-2109.2011.02851.x
|
| [72] | Hatje E, Neuman C, Stevenson H, et al. (2014) Population dynamics of Vibrio and Pseudomonas species isolated from farmed Tasmanian Atlantic salmon (Salmo salar L.): A seasonal study. Microb Ecol 68: 679–687. |
| [73] |
Foulon V, Le Roux F, Lambert C, et al. (2016) Colonization of polystyrene microparticles by Vibrio crassostreae: Light and electron microscopic investigation. Environ Sci Technol 50: 10988–10996. doi: 10.1021/acs.est.6b02720
|
| [74] | Zha H, Jeffs A, Dong Y, et al. (2018) Potential virulence factors of bacteria associated with tail fan necrosis in the spiny lobster, Jasus edwardsii. J Fish Dis 31: 817–828. |
| [75] | Fontanez KM, Eppley JM, Samo TJ, et al. (2015) Microbial community structure and function on sinking particles in the North Pacific Subtropical Gyre. Front Microbiol 6: 469. |
| [76] | Vandecandelaere I, Nercessian O, Segaert E, et al. (2008) Ruegeria scottomollicae sp. nov., isolated from a marine electroactive biofilm. Int J Syst Evol Microbiol 58: 2726–2733. |
| [77] |
Magalhães C, Kiene RP, Buchan A, et al. (2012) A novel inhibitory interaction between dimethylsulfoniopropionate (DMSP) and the denitrification pathway. Biogeochemistry 107: 393–408. doi: 10.1007/s10533-010-9560-0
|
| [78] |
Smedile F, Messina E, La Cono V, et al. (2013) Metagenomic analysis of hadopelagic microbial assemblages thriving at the deepest part of Mediterranean Sea, Matapan-Vavilov Deep. Environ Microbiol 15: 167–182. doi: 10.1111/j.1462-2920.2012.02827.x
|
| [79] |
Luna G (2015) Diversity of marine microbes in a changing Mediterranean Sea. Rend Lincei 26: 49–58. doi: 10.1007/s12210-014-0333-x
|
| [80] |
Pukall R, Päuker O, Buntefuß D, et al. (1999) High sequence diversity of Alteromonas macleodii-related cloned and cellular 16S rDNAs from a Mediterranean seawater mesocosm experiment. FEMS Microbiol Ecol 28: 335–344. doi: 10.1111/j.1574-6941.1999.tb00588.x
|
| [81] |
Garcia-Martinez J, Acinas SG, Massana R, et al. (2002) Prevalence and microdiversity of Alteromonas macleodii-like microorganisms in different oceanic regions. Environ Microbiol 4: 42–50. doi: 10.1046/j.1462-2920.2002.00255.x
|
| [82] |
Ivars-Martinez E, D'Auria G, Rodriguez-Valera F, et al. (2008) Biogeography of the ubiquitous marine bacterium Alteromonas macleodii determined by multilocus sequence analysis. Mol Ecol 17: 4092–4106. doi: 10.1111/j.1365-294X.2008.03883.x
|
| [83] |
Klochko V, Zelena L, Ivoychuk S, et al. (2012) Peculiarities of Alteromonas macleodii strains reflects their deep/surface habitation rather than geographical distribution. J Gen Appl Microbiol 58: 129–135. doi: 10.2323/jgam.58.129
|
| [84] | López-Pérez M, Rodriguez-Valera F, (2014) The family Alteromonadaceae, In: The Prokaryotes: Gammaproteobacteria, 69–92. |
| [85] | Hou S, López-Pérez M, Pfreundt U, et al. (2018) Benefit from decline: The primary transcriptome of Alteromonas macleodii str. Te101 during Trichodesmium demise. ISME J 12: 981–996. |
| [86] |
Lee MD, Walworth NG, Mcparland EL, et al. (2017) The Trichodesmium consortium: Conserved heterotrophic co-occurrence and genomic signatures of potential interactions. ISME J 11: 1813–1824. doi: 10.1038/ismej.2017.49
|
| [87] |
Chi WJ, Lim JH, Park DY, et al. (2013) Isolation and characterization of a novel agar degrading bacterium, Alteromonas macleodii subsp. GNUM08120, from red macroalgae. Korean J Microbiol Biotechnol 41: 8–16. doi: 10.4014/kjmb.1208.08001
|
| [88] |
Neumann AM, Balmonte JP, Berger M, et al. (2015) Different utilization of alginate and other algal polysaccharides by marine Alteromonas macleodii ecotypes. Environ Microbiol 17: 3857–3868. doi: 10.1111/1462-2920.12862
|
| [89] |
Mitulla M, Dinasquet J, Guillemette R, et al. (2016) Response of bacterial communities from California coastal waters to alginate particles and an alginolytic Alteromonas macleodii strain. Environ Microbiol 18: 4369–4377. doi: 10.1111/1462-2920.13314
|
| [90] |
Allers E, Niesner C, Wild C, et al. (2008) Microbes enriched in seawater after addition of coral mucus. Appl Environ Microb 74: 3274–3278. doi: 10.1128/AEM.01870-07
|
| [91] |
O'Connor NJ, Richardson DL (1998) Attachment of barnacle (Balanus amphitrite Darwin) larvae: Responses to bacterial films and extracellular materials. J Exp Mar Biol Ecol 226: 115–129. doi: 10.1016/S0022-0981(97)00242-6
|
| [92] | Goto S, Tada Y, Suzuki K, et al. (2017) Production and reutilization of fluorescent dissolved organic matter by a marine bacterial strain, Alteromonas macleodii. Front Microbiol 8: 507. |
| [93] | Varbanets LD, Avdeeva LV, Borzova NV, et al. (2011) The Black Sea bacteria-producers of hydrolytic enzymes. Mikrobiol Z 73: 9–15. |
| [94] | Kesarcodi-Watson A, Kaspar H, Lategan MJ, et al. (2010) Alteromonas macleodii 0444 and Neptunomonas sp. 0536, two novel probiotics for hatchery-reared Greenshell™ mussel larvae, Perna canaliculus. Aquaculture 309: 49–55. |
| [95] |
Raguénès G, Cambon-Bonavita MA, Lohier JF, et al. (2003) A novel, highly viscous polysaccharide excreted by an Alteromonas isolated from a deep-sea hydrothermal vent shrimp. Curr Microbiol 46: 448–452. doi: 10.1007/s00284-002-3922-3
|
| [96] |
Cambon-Bonavita MA, Raguenes G, Jean J, et al. (2002) A novel polymer produced by a bacterium isolated from a deep-sea hydrothermal vent polychaete annelid. J Appl Microbiol 93: 310–315. doi: 10.1046/j.1365-2672.2002.01689.x
|
| [97] | Khandeparker L, Gaonkar CC, Desai DV (2013) Degradation of barnacle nauplii: Implications to chitin regulation in the marine environment. Biologia 68: 696–706. |
 microbiol-04-03-397-s1.pdf microbiol-04-03-397-s1.pdf |
 |
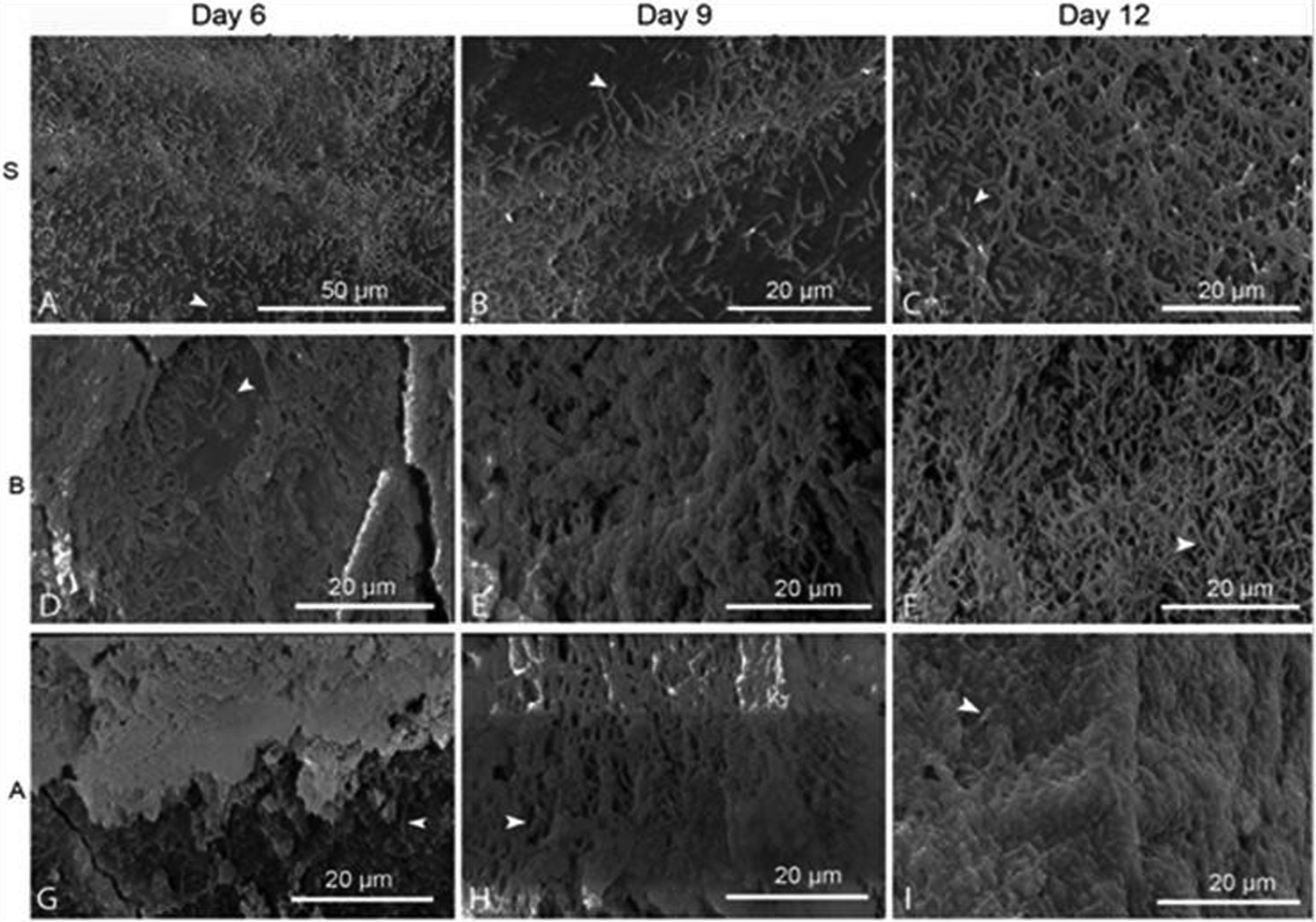
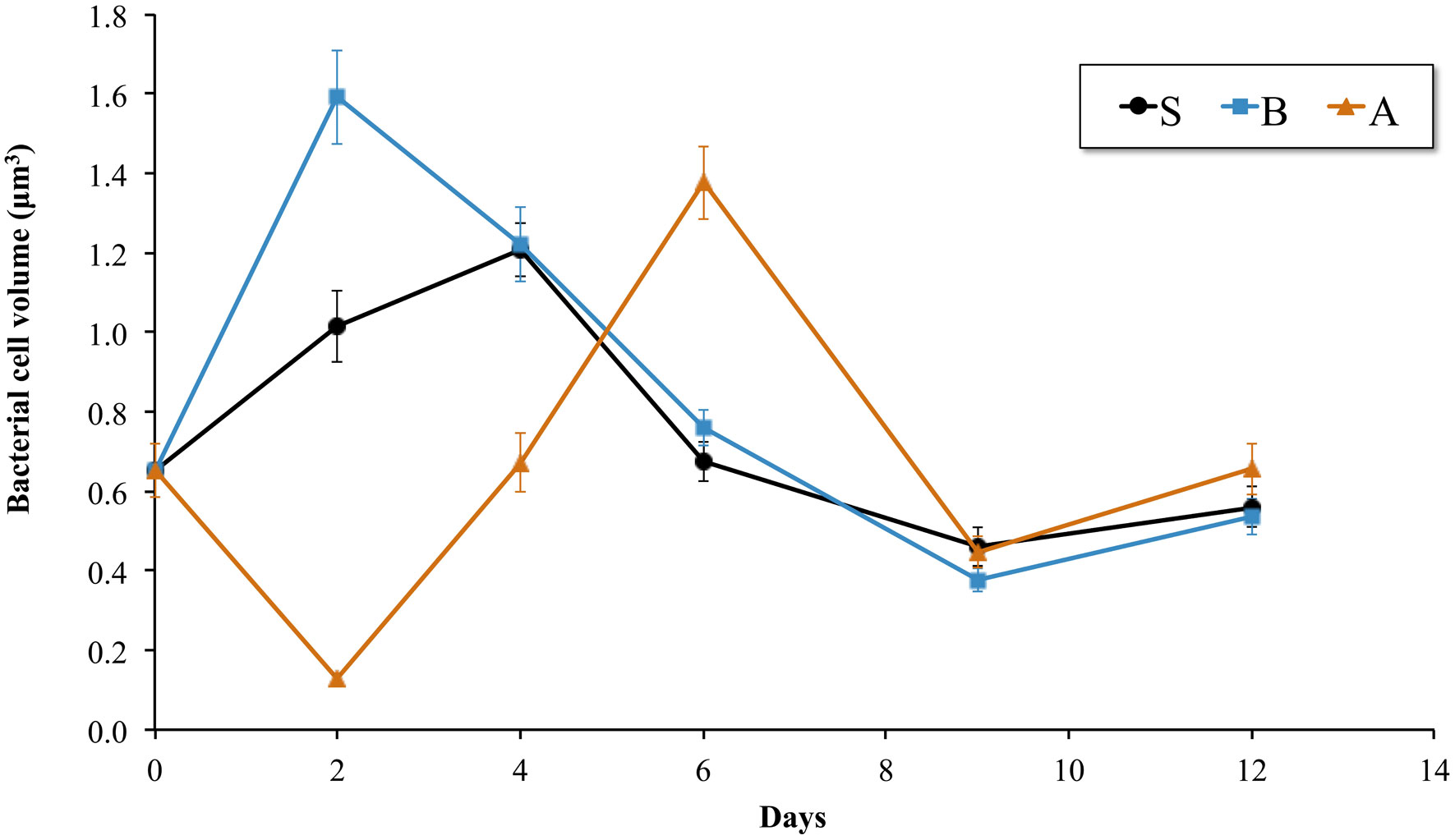
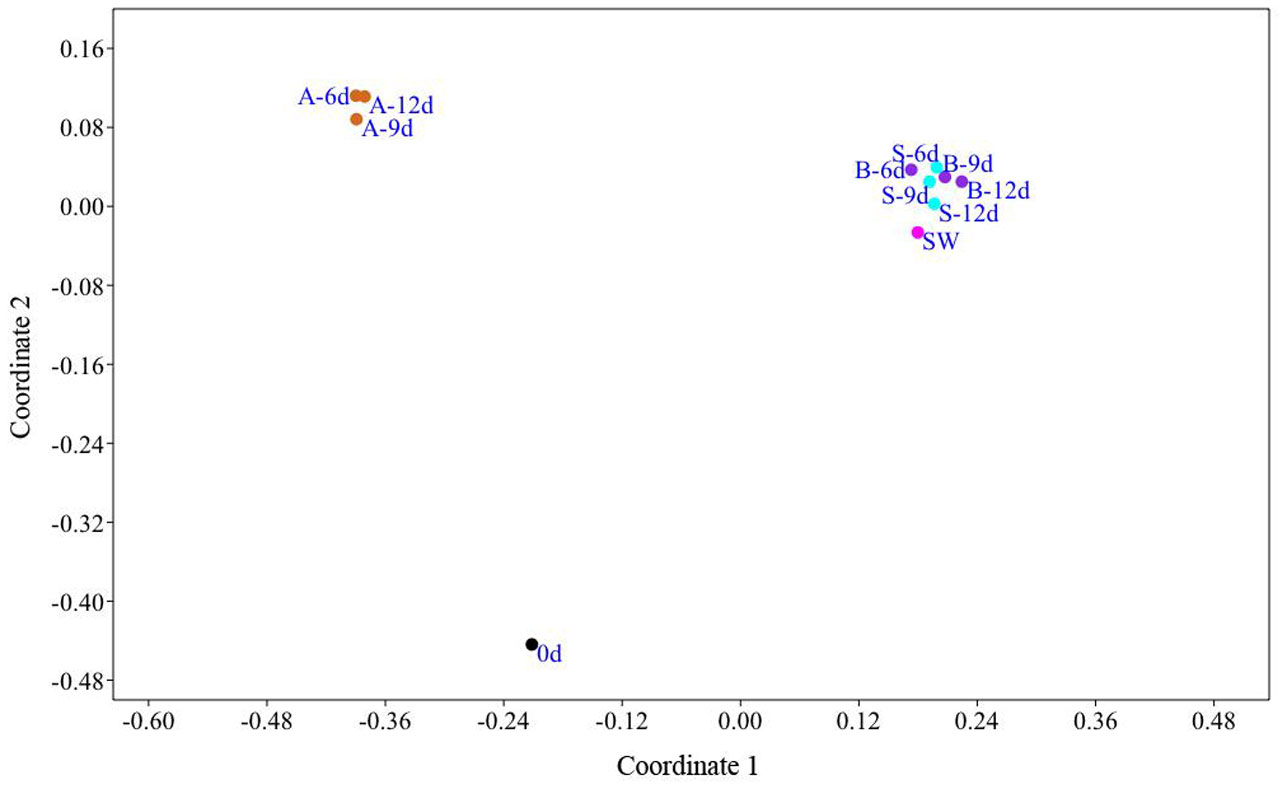
Figures(3) / Tables(1)

Nikolina-Alexandra Xaxiri, Eleni Nikouli, Panagiotis Berillis, Konstantinos Ar. Kormas. Bacterial biofilm development during experimental degradation of Melicertus kerathurus exoskeleton in seawater[J]. AIMS Microbiology, 2018, 4(3): 397-412. doi: 10.3934/microbiol.2018.3.397







 DownLoad:
DownLoad: