Citation: Kamila Tomoko Yuyama, Thaís Souto Paula da Costa Neves, Marina Torquato Memória, Iago Toledo Tartuci, Wolf-Rainer Abraham. Aurantiogliocladin inhibits biofilm formation at subtoxic concentrations[J]. AIMS Microbiology, 2017, 3(1): 50-60. doi: 10.3934/microbiol.2017.1.50

| [1] | Leone M,Dillon LR (2008) Catheter outcomes in home infusion. J Infus Nurs 31: 84–91. |
| [2] | Singh PK, Schaefer AL,Parsek MR, et al. (2000) Quorum-sensing signals indicate that cystic fibrosis lungs are infected with bacterial biofilms. Nature 407: 762–764. |
| [3] |
Hall-Stoodley L,Hu FZ, Gieseke A, et al. (2006) Direct detection of bacterial biofilms on the middle-ear mucosa of children with chronic otitis media. JAMA 296:202–211. doi: 10.1001/jama.296.2.202

|
| [4] |
Rosen Da HT, Stamm WE, Humphrey PA, et al. (2007) Detection of intracellular bacterial communities in human urinary tract infection. PLoS Med 4: e329. doi: 10.1371/journal.pmed.0040329
|
| [5] |
Carron MA, Tran VR, Sugawa C, et al. (2006) Identification of Helicobacter pylori biofilms in human gastric mucosa. J Gastrointest Surg 10:712–717. doi: 10.1016/j.gassur.2005.10.019
|
| [6] |
Hall-Stoodley L, Costerton JW, Stoodley P (2004) Bacterial biofilms: From the natural environment to infectious diseases. Nat Rev Microbiol 2: 95–108. doi: 10.1038/nrmicro821
|
| [7] |
Camilli A, Bassler BL (2006) Bacterial small-molecule signaling pathways. Science 311: 1113–1116. doi: 10.1126/science.1121357
|
| [8] |
Schauder S, Bassler BL (2001) The languages of bacteria. Genes Dev 15:1468–1480. doi: 10.1101/gad.899601
|
| [9] |
Abraham WR (2005) Controlling Gram-negative pathogenic bacteria by interfering with their biofilm formation. Drug Design Rev Online 2: 13–33. doi: 10.2174/1567269053390257
|
| [10] |
Chen X, Schauder S, Potier N, et al. (2002) Structural identification of a bacterial quorum-sensing signal containing boron. Nature 415:545–549. doi: 10.1038/415545a
|
| [11] |
Lyon GJ, Novick RP (2004) Peptide signaling in Staphylococcus aureus and other Gram-positive bacteria. Peptides 25: 1389–1403. doi: 10.1016/j.peptides.2003.11.026
|
| [12] |
Stewart PS, Costerton JW (2001) Antibiotic resistance of bacteria in biofilms. Lancet 358: 135–138. doi: 10.1016/S0140-6736(01)05321-1
|
| [13] | Joly V, Pangon B,Vallois JM, et al. (1987) Value of antibiotic levels in serum and cardiac vegetations for predicting antibacterial effect of ceftriaxone in experimental Escherichia coli endocarditis. Antimicrob Agents Chemother 31:1632–1639. |
| [14] |
Austin DJ, Kristinsson KG, Anderson RM (1999) The relationship between the volume of antimicrobial consumption in human communities and the frequency of resistance. Proc Natl Acad Sci USA 96: 1152–1156. doi: 10.1073/pnas.96.3.1152
|
| [15] | Payne DJ (2008) Microbiology. Desperately seeking new antibiotics. Science 321: 1644–1645. |
| [16] |
Worthington RJ, Richards JJ, Melander C (2014) Non-microbicidal control of bacterial biofilms with small molecules. Anti-Inf Agents 12: 120–138. doi: 10.2174/22113525113119990107
|
| [17] |
Zhu J, Kaufmann GF (2013) Quo vadis quorum quenching? Curr Opin Pharmacol 13: 688–898. doi: 10.1016/j.coph.2013.07.003
|
| [18] |
de Carvalho MP, Türck P, Abraham WR (2015) Secondary metabolites control the associated bacterial communities of saprophytic Basidiomycotina fungi. Microbes Environ 30: 196–198. doi: 10.1264/jsme2.ME14139
|
| [19] |
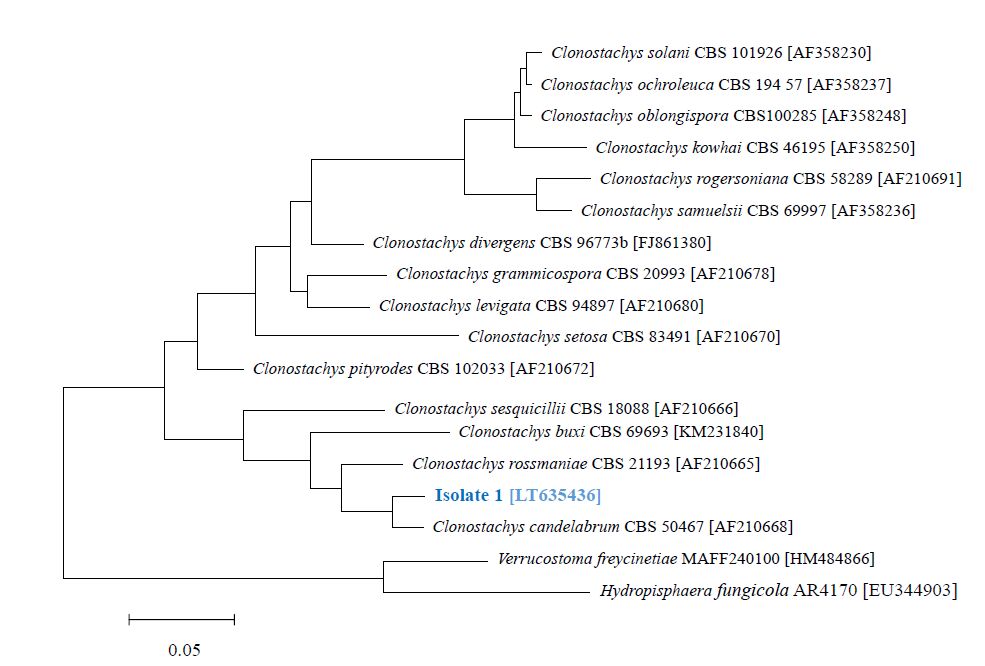
Bruns TD, Fogel R, White TJ, et al. (1990) Amplification and sequencing of DNA from fungal herbarium specimens. Mycologia 82: 175–184. doi: 10.2307/3759846
|
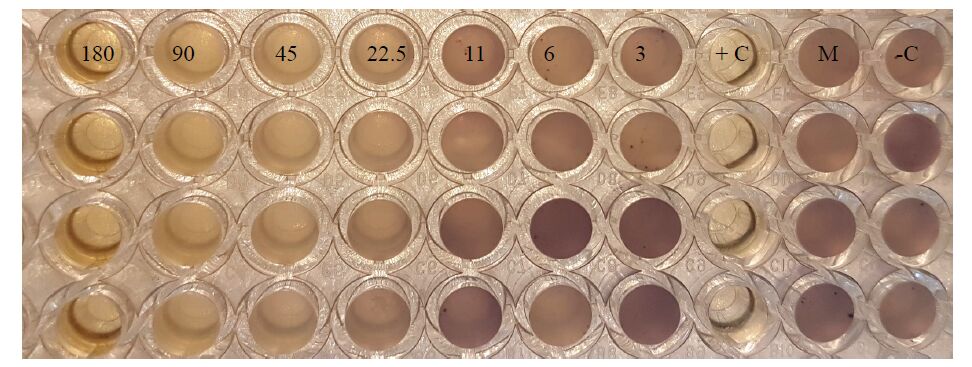
| [20] | O'Toole GA (2011) Microtiter dish biofilm formation assay. J Vis Exp 47: e2437–e2437. |
| [21] | Franke C, Krohn K (2008) Aurantiogliocladin, In: Roth L, Rupp G, Roth collection of natural products data: concise descriptions and spectra , Weinheim, John Wiley & Sons. |
| [22] |
Brian PW, Curtis PJ, Howland SR, et al. (1951) Three new antibiotics from a species of Gliocladium. Experientia 7: 266–267. doi: 10.1007/BF02154546
|
| [23] | Vischer EB (1953) The structures of aurantio-and rubro-gliocladin and gliorosein. J Chem Soc (Resumed) , 815–820. |
| [24] | Bentley R, Lavate WV (1965) Studies on Coenzyme Q. The biosynthesis of aurantiogliocladin and coenzyme Q in molds. J Biol Chem 240: 532–540. |
| [25] |
Pettersson G (1965) New phenolic metabolites from Gliocladium roseum. Acta Chem Scand 19:414–420. doi: 10.3891/acta.chem.scand.19-0414
|
| [26] |
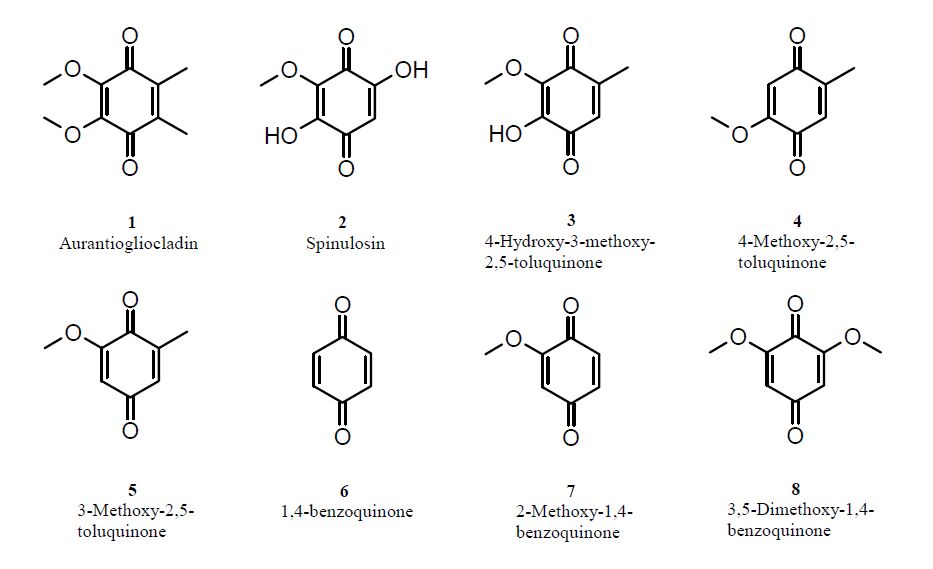
Ayers S, Zink DL, Mohn K, et al. (2010). Anthelmintic constituents of Clonostachys candelabrum. J Antibiotics 63:119–122. doi: 10.1038/ja.2009.131
|
| [27] |
Mozaina K, Cantrell CL, Mims AB, et al. (2008) Activity of 1,4-benzoquinones against Formosan subterranean termites (Coptotermes formosanus ).J Agric Food Chem 56:4021–4026. doi: 10.1021/jf800331r
|
| [28] | Likhovidov VE, Isangalin FS, Naumov AN, et al. (2010) Agents against mosquitos. Russ patent , RU 2391389 C2 20100610. |
| [29] | Klink JW, Dropkin VH, Mitchell JE (1970) Studies on the host-finding mechanisms of Neotylenchus linfordi. J Nematol 2:106–117. |
| [30] |
Birkinshaw JH, Raistrick H (1931) On a new methoxy-dihydroxy-toluquinone produced from glucose by species of Penicillium of the P. spinulosum series.Phil Trans Roy Soc London. B 220: 245–254. doi: 10.1098/rstb.1931.0023
|
| [31] |
Petterson G (1963) Toluquinones from Aspergillus fumigatus. Acta Chem Scand 17:1771–1776. doi: 10.3891/acta.chem.scand.17-1771
|
| [32] | Lauer U, Anke T, Hansske F (1991) Antibiotics from basidiomycetes. XXXVIII. 2-Methoxy-5- methyl-1,4-benzoquinone, a thromboxane A2 receptor antagonist from Lentinus adhaerens. J Antibiot 44:59–65. |
| [33] |
Attygalle AB, Xu SC, Meinwald J, et al. (1993). Defensive secretion of the millipede Floridobolus penneri. J Nat Prod 56:1700–1706. doi: 10.1021/np50100a007
|
| [34] | Nishina A, Uchibori T (1991) Antimicrobial activity of 2, 6-dimethoxy-p-benzoquinone, isolated from thick-stemmed bamboo, its analogs. Agric Biol Chem 55:2395–2398. |
| [35] |
Anchel M, Hervey A, Kavanagh F, et al. (1948) Antibiotic substances from Basidiomycetes III. Coprinus similis and Lentinus degener. Proc Nat Acad Sci 34:498–502. doi: 10.1073/pnas.34.11.498
|
| [36] | Sood RS, Roy K, Reddy GCS, et al. (1982) 3-Methoxy-2, 5-toluquinone from Aspergillus sp. HPL Y-30,212. Fermentation, isolation, characterization and biological properties. J Antibiot 35:985–987. |
| [37] | Oxford AE (1942) Bacteriostatic powers of the methyl ethers of fumigatin and spinulosin and other hydroxy-, methoxy- and hydroxymethoxy-derivatives of toluquinone and benzoquinone. Chem Ind 61:189–192. |
| [38] |
Lana EJ, Carazza F, Takahashi JA (2006) Antibacterial evaluation of 1, 4-benzoquinone derivatives. J Agric Food Chem 54:2053–2056. doi: 10.1021/jf052407z
|
| [39] |
Estrela AB, Abraham W-R (2010) Combining biofilm-controlling compounds and antibiotics as a promising new way to control biofilm infections. Pharmaceuticals 3:1374–1393. doi: 10.3390/ph3051374
|
| [40] |
Wargo MJ, Hogan DA (2006) Fungal-bacterial interactions: a mixed bag of mingling microbes. Curr Opin Microbiol 9: 359–364. doi: 10.1016/j.mib.2006.06.001
|
Figures(3) / Tables(2)

Kamila Tomoko Yuyama, Thaís Souto Paula da Costa Neves, Marina Torquato Memória, Iago Toledo Tartuci, Wolf-Rainer Abraham. Aurantiogliocladin inhibits biofilm formation at subtoxic concentrations[J]. AIMS Microbiology, 2017, 3(1): 50-60. doi: 10.3934/microbiol.2017.1.50







 DownLoad:
DownLoad: