Citation: Ka-man Fong, Shek-yin Au, Ka-lee Lily Chan, Wing-yiu George Ng. Update on management of acute respiratory distress syndrome[J]. AIMS Medical Science, 2018, 5(2): 145-161. doi: 10.3934/medsci.2018.2.145

| [1] |
Wheeler AP, Bernard GR (2007) Acute lung injury and the acute respiratory distress syndrome: A clinical review. Lancet 369: 1553–1564. doi: 10.1016/S0140-6736(07)60604-7

|
| [2] |
Bellani G, Laffey JG, Pham T, et al. (2016) Epidemiology, Patterns of Care, and Mortality for Patients With Acute Respiratory Distress Syndrome in Intensive Care Units in 50 Countries. JAMA 315: 788–800. doi: 10.1001/jama.2016.0291
|
| [3] | Ranieri VM, Rubenfeld GD, Thompson BT, et al. (2012) Acute respiratory distress syndrome: The Berlin Definition. JAMA 307: 2526–2533. |
| [4] |
Bernard GR, Artigas A, Brigham KL, et al. (1994) The American-European consensus conference on ARDS: Definitions, mechanisms, relevant outcomes, and clinical trial coordination. Am J Respir Crit Care Med 149: 818–824. doi: 10.1164/ajrccm.149.3.7509706
|
| [5] |
Meade MO, Cook RJ, Guyatt GH, et al. (2000) Interobserver variation in interpreting chest radiographs for the diagnosis of acute respiratory distress syndrome. Am J Respir Crit Care Med 161: 85–90. doi: 10.1164/ajrccm.161.1.9809003
|
| [6] | Peng JM, Qian CY, Yu XY, et al. (2017) Does training improve diagnostic accuracy and inter-rater agreement in applying the Berlin radiographic definition of acute respiratory distress syndrome? A multicenter prospective study. Crit Care 21: 12. |
| [7] |
Gajic O, Dara SI, Mendez JL, et al. (2004) Ventilator-associated lung injury in patients without acute lung injury at the onset of mechanical ventilation. Crit Care Med 32: 1817–1824. doi: 10.1097/01.CCM.0000133019.52531.30
|
| [8] |
Anonymous (1999) International consensus conferences in intensive care medicine: Ventilator-associated Lung Injury in ARDS. Am J Respir Crit Care Med 160: 2118–2124. doi: 10.1164/ajrccm.160.6.ats16060
|
| [9] |
Pesenti AM (2005) The concept of "baby lung". Intensive Care Med 31: 776–784. doi: 10.1007/s00134-005-2627-z
|
| [10] |
Curley GF, Laffey JG, Zhang H, et al. (2016) Biotrauma and Ventilator-Induced Lung Injury: Clinical Implications. Chest 150: 1109–1117. doi: 10.1016/j.chest.2016.07.019
|
| [11] |
Thompson BT, Chambers RC, Liu KD (2017) Acute Respiratory Distress Syndrome. N Engl J Med 377: 562–572. doi: 10.1056/NEJMra1608077
|
| [12] |
Amato MB, Barbas CS, Medeiros DM, et al. (1998) Effect of a protective-ventilation strategy on mortality in the acute respiratory distress syndrome. N Engl J Med 338: 347–354. doi: 10.1056/NEJM199802053380602
|
| [13] |
Ards N (2000) Ventilation with lower tidal volumes as compared with traditional tidal volumes for acute lung injury and the acute respiratory distress syndrome. N Engl J Med 342: 1301–1308. doi: 10.1056/NEJM200005043421801
|
| [14] |
Walkey AJ, Goligher E, Del SL, et al. (2017) Low Tidal Volume versus Non-Volume-Limited Strategies for Patients with Acute Respiratory Distress Syndrome: A Systematic Review and Meta-Analysis. Ann Am Thorac Soc 14: S271–S279. doi: 10.1513/AnnalsATS.201704-337OT
|
| [15] |
Fan E, Del SL, Goligher EC, et al. (2017) An Official American Thoracic Society/European Society of Intensive Care Medicine/Society of Critical Care Medicine Clinical Practice Guideline: Mechanical Ventilation in Adult Patients with Acute Respiratory Distress Syndrome. Am J Respir Crit Care Med 195: 1253–1263. doi: 10.1164/rccm.201703-0548ST
|
| [16] |
Lang JD, Chumley P, Eiserich JP, et al. (2000) Hypercapnia induces injury to alveolar epithelial cells via a nitric oxide-dependent pathway. Am J Physiol Lung Cell Mol Physiol 279: 994–1002. doi: 10.1152/ajplung.2000.279.5.L994
|
| [17] |
Morimont P, Batchinsky A, Lambermont B (2015) Update on the role of extracorporeal CO 2 removal as an adjunct to mechanical ventilation in ARDS. Crit Care 19: 117. doi: 10.1186/s13054-015-0799-7
|
| [18] | Repessé X, Vieillardbaron A (2017) Hypercapnia during acute respiratory distress syndrome: The tree that hides the forest! J Thorac Dis 9: 1420–1425. |
| [19] |
Nin N, Muriel A, Penuelas O, et al. (2017) Severe hypercapnia and outcome of mechanically ventilated patients with moderate or severe acute respiratory distress syndrome. Intensive Care Med 43: 200–208. doi: 10.1007/s00134-016-4611-1
|
| [20] | Barnes T, Zochios V, Parhar K (2017) Re-examining permissive hypercapnia in ARDS: A narrative review. Chest. |
| [21] | Sahetya SK, Goligher EC, Brower RG (2017) Fifty Years of Research in ARDS. Setting Positive End-Expiratory Pressure in Acute Respiratory Distress Syndrome. Am J Respir Crit Care Med 195: 1429–1438. |
| [22] |
Brower RG, Lanken PN, MacIntyre N, et al. (2004) Higher versus lower positive end-expiratory pressures in patients with the acute respiratory distress syndrome. N Engl J Med 351: 327–336. doi: 10.1056/NEJMoa032193
|
| [23] |
Meade MO, Cook DJ, Guyatt GH, et al. (2008) Ventilation strategy using low tidal volumes, recruitment maneuvers, and high positive end-expiratory pressure for acute lung injury and acute respiratory distress syndrome: A randomized controlled trial. JAMA 299: 637–645. doi: 10.1001/jama.299.6.637
|
| [24] |
Mercat A, Richard JC, Vielle B, et al. (2008) Positive end-expiratory pressure setting in adults with acute lung injury and acute respiratory distress syndrome: A randomized controlled trial. JAMA 299: 646–655. doi: 10.1001/jama.299.6.646
|
| [25] | Santa CR, Rojas JI, Nervi R, et al. (2013) High versus low positive end-expiratory pressure (PEEP) levels for mechanically ventilated adult patients with acute lung injury and acute respiratory distress syndrome. Cochrane Database Syst Rev 6: CD009098. |
| [26] |
Talmor D, Sarge T, Malhotra A, et al. (2008) Mechanical ventilation guided by esophageal pressure in acute lung injury. N Engl J Med 359: 2095–2104. doi: 10.1056/NEJMoa0708638
|
| [27] |
Fish E, Novack V, Bannergoodspeed VM, et al. (2014) The Esophageal Pressure-Guided Ventilation 2 (EPVent2) trial protocol: A multicentre, randomised clinical trial of mechanical ventilation guided by transpulmonary pressure. BMJ Open 4: e006356. doi: 10.1136/bmjopen-2014-006356
|
| [28] |
Kacmarek RM, Villar J, Sulemanji D, et al. (2016) Open Lung Approach for the Acute Respiratory Distress Syndrome: A Pilot, Randomized Controlled Trial. Crit Care Med 44: 32–42. doi: 10.1097/CCM.0000000000001383
|
| [29] | Writing Group for the Alveolar Recruitment for Acute Respiratory Distress Syndrome Trial I, Cavalcanti AB, Suzumura EA, et al. (2017) Effect of Lung Recruitment and Titrated Positive End-Expiratory Pressure (PEEP) vs. Low PEEP on Mortality in Patients With Acute Respiratory Distress Syndrome: A Randomized Clinical Trial. JAMA 318: 1335–1345. |
| [30] |
Luecke T, Corradi F, Pelosi P (2012) Lung imaging for titration of mechanical ventilation. Curr Opin Anaesthesiol 25: 131–140. doi: 10.1097/ACO.0b013e32835003fb
|
| [31] |
Jabaudon M, Godet T, Futier E, et al. (2017) Rationale, study design and analysis plan of the lung imaging morphology for ventilator settings in acute respiratory distress syndrome study (LIVE study): Study protocol for a randomised controlled trial. Anaesthesia Crit Care Pain Med 36: 301–306. doi: 10.1016/j.accpm.2017.02.006
|
| [32] |
Bugedo G, Retamal J, Bruhn A (2017) Driving pressure: A marker of severity, a safety limit, or a goal for mechanical ventilation? Crit Care 21: 199. doi: 10.1186/s13054-017-1779-x
|
| [33] |
Amato MB, Meade MO, Slutsky AS, et al. (2015) Driving pressure and survival in the acute respiratory distress syndrome. N Engl J Med 372: 747–755. doi: 10.1056/NEJMsa1410639
|
| [34] |
Estenssoro E, Dubin A, Laffaire E, et al. (2002) Incidence, clinical course, and outcome in 217 patients with acute respiratory distress syndrome. Crit Care Med 30: 2450–2456. doi: 10.1097/00003246-200211000-00008
|
| [35] |
Papazian L, Forel JM, Gacouin A, et al. (2010) Neuromuscular blockers in early acute respiratory distress syndrome. N Engl J Med 363: 1107–1116. doi: 10.1056/NEJMoa1005372
|
| [36] |
Guerin C, Reignier J, Richard JC, et al. (2013) Prone positioning in severe acute respiratory distress syndrome. N Engl J Med 368: 2159–2168. doi: 10.1056/NEJMoa1214103
|
| [37] |
Kassis EB, Loring SH, Talmor D (2016) Mortality and pulmonary mechanics in relation to respiratory system and transpulmonary driving pressures in ARDS. Intensive Care Med 42: 1206–1213. doi: 10.1007/s00134-016-4403-7
|
| [38] |
Ferguson ND, Cook DJ, Guyatt GH, et al. (2013) High-frequency oscillation in early acute respiratory distress syndrome. N Engl J Med 368: 795–805. doi: 10.1056/NEJMoa1215554
|
| [39] |
Young D, Lamb SE, Shah S, et al. (2013) High-frequency oscillation for acute respiratory distress syndrome. N Engl J Med 368: 806–813. doi: 10.1056/NEJMoa1215716
|
| [40] |
Davies SW, Leonard KL, Falls RK, et al. (2015) Lung protective ventilation (ARDSNet) versus airway pressure release ventilation: Ventilatory management in a combined model of acute lung and brain injury. Trauma Acute Care Surg 78: 240–249. doi: 10.1097/TA.0000000000000518
|
| [41] |
Mireles-Cabodevila E, Kacmarek RM (2016) Should Airway Pressure Release Ventilation Be the Primary Mode in ARDS? Respir Care 61: 761–773. doi: 10.4187/respcare.04653
|
| [42] |
Zhou Y, Jin X, Lv Y, et al. (2017) Early application of airway pressure release ventilation may reduce the duration of mechanical ventilation in acute respiratory distress syndrome. Intensive Care Med 43: 1648–1659. doi: 10.1007/s00134-017-4912-z
|
| [43] |
Bellani G, Laffey JG, Pham T, et al. (2017) Noninvasive Ventilation of Patients with Acute Respiratory Distress Syndrome. Insights from the LUNG SAFE Study. Am J Respir Crit Care Med 195: 67–77. doi: 10.1164/rccm.201606-1306OC
|
| [44] | Patel BK, Wolfe KS, Pohlman AS, et al. (2016) Effect of Noninvasive Ventilation Delivered by Helmet vs. Face Mask on the Rate of Endotracheal Intubation in Patients With Acute Respiratory Distress Syndrome: A Randomized Clinical Trial. JAMA 315: 2435–2441. |
| [45] |
Hernandez G, Roca O, Colinas L (2017) High-flow nasal cannula support therapy: New insights and improving performance. Crit Care 21: 62. doi: 10.1186/s13054-017-1640-2
|
| [46] |
Parke RL, Mcguinness SP (2013) Pressures delivered by nasal high flow oxygen during all phases of the respiratory cycle. Respir Care 58: 1621–1624. doi: 10.4187/respcare.02358
|
| [47] | Drake MG (2017) High Flow Nasal Cannula Oxygen in Adults: An Evidence-Based Assessment. Ann Am Thorac Soc 15: 145–155. |
| [48] |
Frat JP, Thille AW, Mercat A, et al. (2015) High-flow oxygen through nasal cannula in acute hypoxemic respiratory failure. N Engl J Med 372: 2185–2196. doi: 10.1056/NEJMoa1503326
|
| [49] |
Kang BJ, Koh Y, Lim CM, et al. (2015) Failure of high-flow nasal cannula therapy may delay intubation and increase mortality. Intensive Care Med 41: 623–632. doi: 10.1007/s00134-015-3693-5
|
| [50] | Ni YN, Luo J, Yu H, et al. (2017) Can High-flow Nasal Cannula Reduce the Rate of Endotracheal Intubation in Adult Patients With Acute Respiratory Failure Compared With Conventional Oxygen Therapy and Noninvasive Positive Pressure Ventilation?: A Systematic Review and Meta-analysis. Chest 151: 764–775. |
| [51] |
Marik PE, Kaufman D (1996) The effects of neuromuscular paralysis on systemic and splanchnic oxygen utilization in mechanically ventilated patients. Chest 109: 1038–1042. doi: 10.1378/chest.109.4.1038
|
| [52] |
Kaisers U, Busch T, Deja M, et al. (2003) Selective pulmonary vasodilation in acute respiratory distress syndrome. Crit Care Med 31: S337–S342. doi: 10.1097/01.CCM.0000057913.45273.1A
|
| [53] |
Griffiths MJ, Evans TW (2005) Inhaled nitric oxide therapy in adults. N Engl J Med 353: 2683–2695. doi: 10.1056/NEJMra051884
|
| [54] |
Markewitz BA, Michael JR (2000) Inhaled nitric oxide in adults with the acute respiratory distress syndrome. Respir Med 94: 1023–1028. doi: 10.1053/rmed.2000.0928
|
| [55] | Gebistorf F, Karam O, Wetterslev J, et al. (2016) Inhaled nitric oxide for acute respiratory distress syndrome (ARDS) in children and adults. Cochrane Database Syst Rev 6: CD002787. |
| [56] |
Fuller BM, Mohr NM, Skrupky L, et al. (2015) The use of inhaled prostaglandins in patients with ARDS: A systematic review and meta-analysis. Chest 147: 1510–1522. doi: 10.1378/chest.14-3161
|
| [57] |
Bassford CR, Thickett DR, Perkins GD (2012) The rise and fall of beta-agonists in the treatment of ARDS. Crit Care 16: 208. doi: 10.1186/cc11221
|
| [58] |
National Heart, Lung, and Blood Institute Acute Respiratory Distress Syndrome (ARDS) Clinical Trials Network, Matthay MA, et al. (2011) Randomized, placebo-controlled clinical trial of an aerosolized beta(2)-agonist for treatment of acute lung injury. Am J Respir Crit Care Med 184: 561–568. doi: 10.1164/rccm.201012-2090OC
|
| [59] |
Gao SF, Perkins GD, Gates S, et al. (2012) Effect of intravenous beta-2 agonist treatment on clinical outcomes in acute respiratory distress syndrome (BALTI-2): A multicentre, randomised controlled trial. Lancet 379: 229–235. doi: 10.1016/S0140-6736(11)61623-1
|
| [60] |
Lai-Fook SJ, Rodarte JR (1991) Pleural pressure distribution and its relationship to lung volume and interstitial pressure. J Appl Physiol 70: 967–978. doi: 10.1152/jappl.1991.70.3.967
|
| [61] |
Pelosi P, D'Andrea L, Vitale G, et al. (1994) Vertical gradient of regional lung inflation in adult respiratory distress syndrome. Am J Respir Crit Care Med 149: 8–13. doi: 10.1164/ajrccm.149.1.8111603
|
| [62] |
Tawhai MH, Nash MP, Lin CL, et al. (2009) Supine and prone differences in regional lung density and pleural pressure gradients in the human lung with constant shape. J Appl Physiol 107: 912–920. doi: 10.1152/japplphysiol.00324.2009
|
| [63] |
Malbouisson LM, Busch CJ, Puybasset L, et al. (2000) Role of the heart in the loss of aeration characterizing lower lobes in acute respiratory distress syndrome. CT Scan ARDS Study Group. Am J Respir Crit Care Med 161: 2005–2012. doi: 10.1164/ajrccm.161.6.9907067
|
| [64] |
Scholten EL, Beitler JR, Prisk GK, et al. (2017) Treatment of ARDS With Prone Positioning. Chest 151: 215–224. doi: 10.1016/j.chest.2016.06.032
|
| [65] |
Mutoh T, Guest RJ, Lamm WJ, et al. (1992) Prone position alters the effect of volume overload on regional pleural pressures and improves hypoxemia in pigs in vivo. Am Rev Respir Dis 146: 300–306. doi: 10.1164/ajrccm/146.2.300
|
| [66] |
Lamm WJ, Graham MM, Albert RK (1994) Mechanism by which the prone position improves oxygenation in acute lung injury. Am J Respir Crit Care Med 150: 184–193. doi: 10.1164/ajrccm.150.1.8025748
|
| [67] | Munshi L, Del LS, Adhikari N, et al. (2017) Prone Position for Acute Respiratory Distress Syndrome. A Systematic Review and Meta-Analysis. Ann Am Thorac Soc 14: S280–S288. |
| [68] |
Peek GJ, Mugford M, Tiruvoipati R, et al. (2009) Efficacy and economic assessment of conventional ventilatory support versus extracorporeal membrane oxygenation for severe adult respiratory failure (CESAR): A multicentre randomised controlled trial. Lancet 374: 1351–1363. doi: 10.1016/S0140-6736(09)61069-2
|
| [69] |
Dembinski R, Hochhausen N, Terbeck S, et al. (2007) Pumpless extracorporeal lung assist for protective mechanical ventilation in experimental lung injury. Crit Care Med 35: 2359–2366. doi: 10.1097/01.CCM.0000281857.87354.A5
|
| [70] |
Schmidt M, Stewart C, Bailey M, et al. (2015) Mechanical ventilation management during extracorporeal membrane oxygenation for acute respiratory distress syndrome: A retrospective international multicenter study. Crit Care Med 43: 654–664. doi: 10.1097/CCM.0000000000000753
|
| [71] |
Neto AS, Schmidt M, Azevedo LCP, et al. (2016) Associations between ventilator settings during extracorporeal membrane oxygenation for refractory hypoxemia and outcome in patients with acute respiratory distress syndrome: A pooled individual patient data analysis: Mechanical ventilation during ECMO. Intensive Care Med 42: 1672–1684. doi: 10.1007/s00134-016-4507-0
|
| [72] |
Tillmann BW, Klingel ML, Iansavichene AE, et al. (2017) Extracorporeal membrane oxygenation (ECMO) as a treatment strategy for severe acute respiratory distress syndrome (ARDS) in the low tidal volume era: A systematic review. J Crit Care 41: 64–71. doi: 10.1016/j.jcrc.2017.04.041
|
| [73] |
Bizzarro MJ, Conrad SA, Kaufman DA, et al. (2011) Infections acquired during extracorporeal membrane oxygenation in neonates, children, and adults. Pediatr Crit Care Med 12: 277–281. doi: 10.1097/PCC.0b013e3181e28894
|
| [74] |
Paden ML, Conrad SA, Rycus PT, et al. (2013) Extracorporeal Life Support Organization Registry Report 2012. ASAIO J 59: 202–210. doi: 10.1097/MAT.0b013e3182904a52
|
| [75] | Mishra V, Svennevig JL, Bugge JF, et al. (2010) Cost of extracorporeal membrane oxygenation: Evidence from the Rikshospitalet University Hospital, Oslo, Norway. Eur J Cardiothorac Surg 37: 339–342. |
| [76] |
Muller T, Lubnow M, Philipp A, et al. (2009) Extracorporeal pumpless interventional lung assist in clinical practice: Determinants of efficacy. Eur Respir J 33: 551–558. doi: 10.1183/09031936.00123608
|
| [77] |
Bein T, Aubron C, Papazian L (2017) Focus on ECMO and ECCO2R in ARDS patients. Intensive Care Med 43: 1424–1426. doi: 10.1007/s00134-017-4882-1
|
| [78] |
Bein T, Weber-Carstens S, Goldmann A, et al. (2013) Lower tidal volume strategy (approximately 3 ml/kg) combined with extracorporeal CO2 removal versus "conventional" protective ventilation (6 ml/kg) in severe ARDS: The prospective randomized Xtravent-study. Intensive Care Med 39: 847–856. doi: 10.1007/s00134-012-2787-6
|
| [79] |
Fitzgerald M, Millar J, Blackwood B, et al. (2014) Extracorporeal carbon dioxide removal for patients with acute respiratory failure secondary to the acute respiratory distress syndrome: A systematic review. Crit Care 18: 222. doi: 10.1186/cc13875
|
| [80] |
Calfee CS, Delucchi K, Parsons PE, et al. (2014) Subphenotypes in acute respiratory distress syndrome: Latent class analysis of data from two randomised controlled trials. Lancet Respir Med 2: 611–620. doi: 10.1016/S2213-2600(14)70097-9
|
| [81] | Famous KR, Delucchi K, Ware LB, et al. (2017) Acute Respiratory Distress Syndrome Subphenotypes Respond Differently to Randomized Fluid Management Strategy. Am J Respir Crit Care Med 195: 331–338. |
| [82] |
Hough CL (2014) Steroids for acute respiratory distress syndrome? Clin Chest Med 35: 781–795. doi: 10.1016/j.ccm.2014.08.014
|
| [83] |
Meduri GU, Bridges L, Shih MC, et al. (2016) Prolonged glucocorticoid treatment is associated with improved ARDS outcomes: Analysis of individual patients' data from four randomized trials and trial-level meta-analysis of the updated literature. Intensive Care Med 42: 829–840. doi: 10.1007/s00134-015-4095-4
|
| [84] | Network TA (2000) Ketoconazole for early treatment of acute lung injury and acute respiratory distress syndrome: A randomized controlled trial. The ARDS Network. JAMA 283: 1995–2002. |
| [85] |
Xiong B, Wang C, Tan J, et al. (2016) Statins for the prevention and treatment of acute lung injury and acute respiratory distress syndrome: A systematic review and meta-analysis. Respirology 21: 1026–1033. doi: 10.1111/resp.12820
|
| [86] |
Sabater J, Masclans JR, Sacanell J, et al. (2008) Effects on hemodynamics and gas exchange of omega-3 fatty acid-enriched lipid emulsion in acute respiratory distress syndrome (ARDS): A prospective, randomized, double-blind, parallel group study. Lipids Health Dis 7: 39. doi: 10.1186/1476-511X-7-39
|
| [87] |
Raghavendran K, Willson D, Notter RH (2011) Surfactant therapy for acute lung injury and acute respiratory distress syndrome. Crit Care Clin 27: 525–559. doi: 10.1016/j.ccc.2011.04.005
|
| [88] |
Shah FA, Girard TD, Yende S (2017) Limiting sedation for patients with acute respiratory distress syndrome-time to wake up. Curr Opin Crit Care 23: 45–51. doi: 10.1097/MCC.0000000000000382
|
| [89] |
National Heart L, Wheeler AP, Wiedemann HP, et al. (2006) Comparison of two fluid-management strategies in acute lung injury. N Engl J Med 354: 2564–2575. doi: 10.1056/NEJMoa062200
|
| [90] |
Jozwiak M, Silva S, Persichini R, et al. (2013) Extravascular lung water is an independent prognostic factor in patients with acute respiratory distress syndrome. Crit Care Med 41: 472–480. doi: 10.1097/CCM.0b013e31826ab377
|
| [91] |
Krzak A, Pleva M, Napolitano LM (2011) Nutrition therapy for ALI and ARDS. Crit Care Clin 27: 647–659. doi: 10.1016/j.ccc.2011.05.004
|
| [92] |
Investigators NS, Finfer S, Chittock DR, et al. (2009) Intensive versus conventional glucose control in critically ill patients. N Engl J Med 360: 1283–1297. doi: 10.1056/NEJMoa0810625
|
| [93] |
Kortebein P (2009) Rehabilitation for hospital-associated deconditioning. Am J Phys Med Rehabil 88: 66–77. doi: 10.1097/PHM.0b013e3181838f70
|
| [94] |
Bailey P, Thomsen GE, Spuhler VJ, et al. (2007) Early activity is feasible and safe in respiratory failure patients. Crit Care Med 35: 139–145. doi: 10.1097/01.CCM.0000251130.69568.87
|
| [95] |
Morris PE, Berry MJ, Files DC, et al. (2016) Standardized Rehabilitation and Hospital Length of Stay Among Patients With Acute Respiratory Failure: A Randomized Clinical Trial. JAMA 315: 2694–2702. doi: 10.1001/jama.2016.7201
|
| [96] | Jabaudon M, Blondonnet R, Audard J, et al. (2017) Recent directions in personalised acute respiratory distress syndrome medicine. Anaesth Crit Care Pain Med. |
| [97] | Shankar-Hari M, Mcauley DF (2017) Acute Respiratory Distress Syndrome Phenotypes and Identifying Treatable Traits. The Dawn of Personalized Medicine for ARDS. Am J Respir Crit Care Med 195: 280–281. |
| [98] |
Wilson JG, Liu KD, Zhuo H, et al. (2015) Mesenchymal stem (stromal) cells for treatment of ARDS: A phase 1 clinical trial. Lancet Respir Med 3: 24–32. doi: 10.1016/S2213-2600(14)70291-7
|
| [99] | Laffey JG, Matthay MA (2017) Fifty Years of Research in ARDS. Cell-based Therapy for Acute Respiratory Distress Syndrome. Biology and Potential Therapeutic Value. Am J Respir Crit Care Med 196: 266–273. |
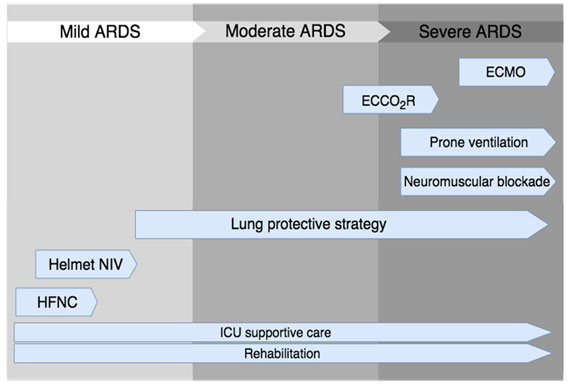
Figures(1) / Tables(2)

Ka-man Fong, Shek-yin Au, Ka-lee Lily Chan, Wing-yiu George Ng. Update on management of acute respiratory distress syndrome[J]. AIMS Medical Science, 2018, 5(2): 145-161. doi: 10.3934/medsci.2018.2.145







 DownLoad:
DownLoad: