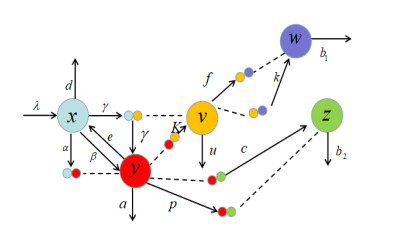
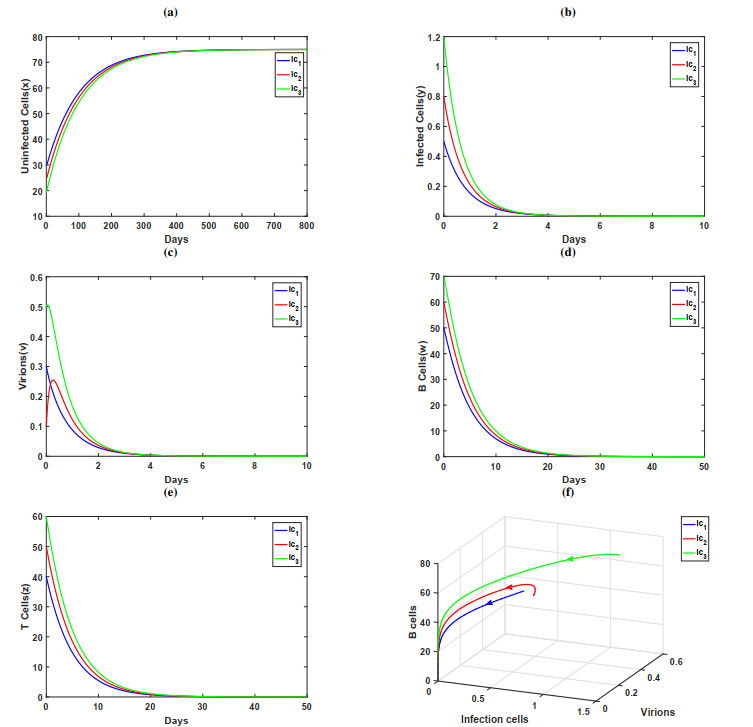
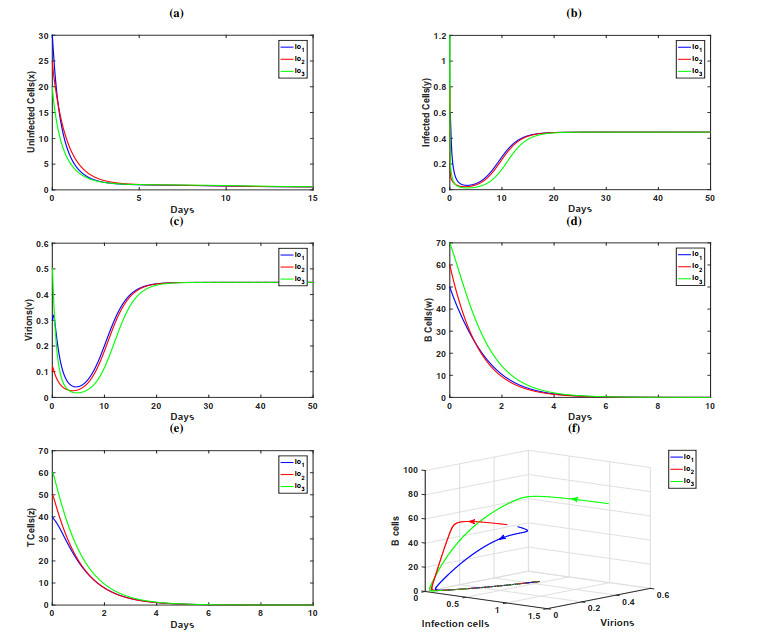
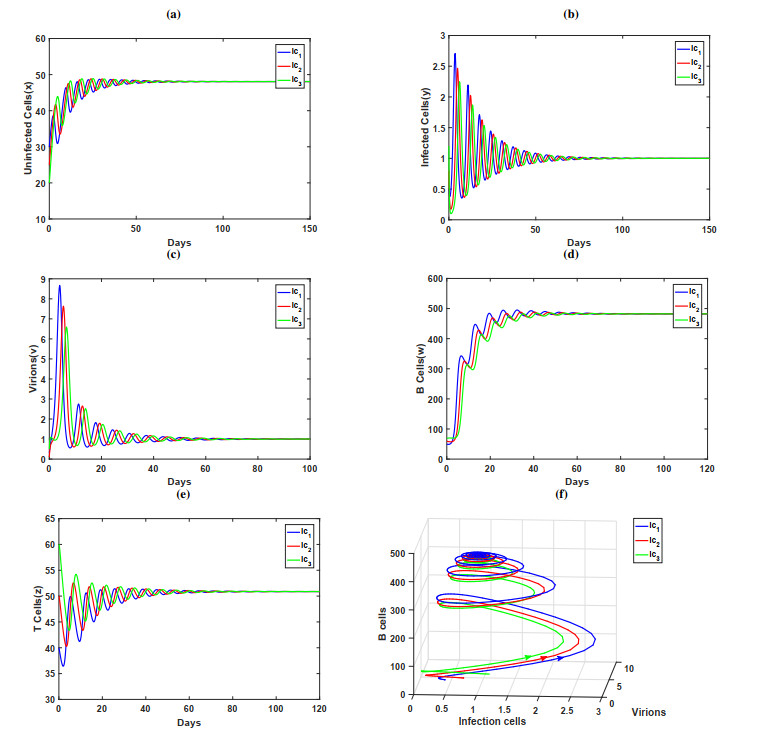
In this paper, a dynamic HIV model with cell-to-cell transmission, two immune responses, and induced apoptosis is proposed and studied. First, the non-negativity and boundedness of the solutions of the model are given, and then the exact expression of the basic reproduction number $ R_{0} $ is obtained by using the next generation matrix method. Second, criteria are obtained for the local stability of the disease-free equilibrium, immune response-free equilibrium, and the infected equilibrium with both humoral and cellular immune responses. Furthermore, the threshold conditions are also derived for the global asymptotic stability of the disease-free equilibrium, immune response-free equilibrium, and the infected equilibrium with both humoral and cellular immune responses by constructing the suitable Lyapunov function. Finally, some numerical simulations are conducted to verify the theoretical results; the numerical simulation results show that the increase of apoptosis rate had a positive role in the control of viral infection.
Citation: Ru Meng, Yantao Luo, Tingting Zheng. Stability analysis for a HIV model with cell-to-cell transmission, two immune responses and induced apoptosis[J]. AIMS Mathematics, 2024, 9(6): 14786-14806. doi: 10.3934/math.2024719
In this paper, a dynamic HIV model with cell-to-cell transmission, two immune responses, and induced apoptosis is proposed and studied. First, the non-negativity and boundedness of the solutions of the model are given, and then the exact expression of the basic reproduction number $ R_{0} $ is obtained by using the next generation matrix method. Second, criteria are obtained for the local stability of the disease-free equilibrium, immune response-free equilibrium, and the infected equilibrium with both humoral and cellular immune responses. Furthermore, the threshold conditions are also derived for the global asymptotic stability of the disease-free equilibrium, immune response-free equilibrium, and the infected equilibrium with both humoral and cellular immune responses by constructing the suitable Lyapunov function. Finally, some numerical simulations are conducted to verify the theoretical results; the numerical simulation results show that the increase of apoptosis rate had a positive role in the control of viral infection.

| [1] | Global HIV and AIDS statistics–Fact sheet, 2023. Available from: https://www.unaids.org/en/resources/fact-sheet. |
| [2] |
M. A. Nowak, C. R. M. Bangham, Population dynamics of immune responses to persistent viruses, Science, 272 (1996), 74–79. http://doi.org/10.1126/science.272.5258.74 doi: 10.1126/science.272.5258.74

|
| [3] |
S. Gummuluru, C. M. Kinsey, M. Emerman, An in vitro rapid-turnover assay for human immunodeficiency virus type 1 replication selects for cell-to-cell spread of virus, J. Virol., 74 (2000), 10882–10891. http://doi.org/10.1128/jvi.74.23.10882-10891.2000 doi: 10.1128/jvi.74.23.10882-10891.2000
|
| [4] |
S. Iwami, J. S. Takeuchi, S. Nakaoka, F. Mammano, F. Clavel, H. Inaba, et al., Cell-to-cell infection by HIV contributes over half of virus infection, Elife, 4 (2015), e08150. http://doi.org/10.7554/eLife.08150.001 doi: 10.7554/eLife.08150.001
|
| [5] |
A. Imle, P. Kumberger, N. D. Schnellbacher, J. Fehr, P. Carrillo-Bustamante, J. Ales, et al., Experimental and computational analyses reveal that environmental restrictions shape HIV-1 spread in 3D cultures, Nat. Commun., 10 (2019), 2144. http://doi.org/10.1038/s41467-019-09879-3 doi: 10.1038/s41467-019-09879-3
|
| [6] |
N. L. Komarova, D. Anghelina, I. Voznesensky, B. Trinité, D. N. Levy, D. Wodarz, Relative contribution of free-virus and synaptic transmission to the spread of HIV-1 through target cell populations, Biol. Lett., 9 (2013), 20121049. http://doi.org/10.1098/rsbl.2012.1049 doi: 10.1098/rsbl.2012.1049
|
| [7] |
M. Huber, A. Trkola, Humoral immunity to HIV-1: neutralization and beyond, J. Intern. Med., 262 (2007), 5–25. http://doi.org/10.1111/j.1365-2796.2007.01819.x doi: 10.1111/j.1365-2796.2007.01819.x
|
| [8] |
W. Hubner, G. P. McNerney, P. Chen, B. M. Dale, R. E. Gordon, F. Y. S. Chuang, et al., Quantitative 3D video microscopy of HIV transfer across T cell virological synapses, Science, 323 (2009), 1743–1747. http://doi.org/10.1126/science.1167525 doi: 10.1126/science.1167525
|
| [9] |
T. Guo, Z. P. Qiu, L. B. Rong, Analysis of an HIV model with immune responses and cell-to-cell transmission, Bull. Malays. Math. Sci. Soc., 43 (2020), 581–607. http://doi.org/10.1007/s40840-018-0699-5 doi: 10.1007/s40840-018-0699-5
|
| [10] |
Y. Wang, J. Liu, X. H. Zhang, J. M. Heffernan, An HIV stochastic model with cell-to-cell infection, B-cell immune response and distributed delay, J. Math. Biol., 86 (2023), 35. http://doi.org/10.1007/s00285-022-01863-8 doi: 10.1007/s00285-022-01863-8
|
| [11] |
J. L. Wang, J. Y. Lang, X. F. Zou, Analysis of an age structured HIV infection model with virus-to-cell infection and cell-to-cell transmission, Nonlinear Anal.-Real., 34 (2017), 75–96 http://doi.org/10.1016/j.nonrwa.2016.08.001 doi: 10.1016/j.nonrwa.2016.08.001
|
| [12] |
Y. Wang, M. M. Lu, D. Q. Jiang, Dynamic behavior of a general stochastic HIV model with virus-to-cell infection, cell-to-cell transmission, immune response and distributed delays, J. Nonlinear Sci., 33 (2023), 97. http://doi.org/10.1007/s00332-023-09955-5 doi: 10.1007/s00332-023-09955-5
|
| [13] |
A. J. McMichael, S. L. Rowland-Jones, Cellular immune responses to HIV, Nature, 410 (2001), 980–987. http://doi.org/10.1038/35073658 doi: 10.1038/35073658
|
| [14] | W. E. Paul, Fundamental immunology, Philadelphia: Lippincott Williams Wilkins, 2012. |
| [15] |
S. F. Wang, D. Y. Zou, Global stability of in-host viral models with humoral immunity and intracellular delays, Appl. Math. Model., 36 (2012), 1313–1322. http://doi.org/10.1016/j.apm.2011.07.086 doi: 10.1016/j.apm.2011.07.086
|
| [16] |
Y. T. Luo, L. Zhang, T. T. Zheng, Z. D. Teng, Analysis of a diffusive virus infection model with humoral immunity, cell-to-cell transmission and nonlinear incidence, Physica A, 535 (2019), 122415. http://doi.org/10.1016/j.physa.2019.122415 doi: 10.1016/j.physa.2019.122415
|
| [17] |
A. M. Elaiw, A. A. Raezah, K. Hattaf, Stability of HIV-1 infection with saturated virus-target and infected-target incidences and CTL immune response, Int. J. Biomath., 10 (2017), 1750070. http://doi.org/10.1142/S179352451750070X doi: 10.1142/S179352451750070X
|
| [18] |
Z. R. Zhu, R. C. Wu, Y. Yang, Y. C. Xu, Modelling HIV dynamics with cell-to-cell transmission and CTL response, Math. Method. Appl. Sci., 46 (2023), 6506–6528. http://doi.org/10.1002/mma.8921 doi: 10.1002/mma.8921
|
| [19] |
J. F. R. Kerr, A. H. Wyllie, A. R. Currie, Apoptosis: A basic biological phenomenon with wideranging implications in tissue kinetics, Br. J. Cancer, 26 (1972), 239–257. http://doi.org/10.1038/bjc.1972.33 doi: 10.1038/bjc.1972.33
|
| [20] |
N. Selliah, T. H. Finkel, Biochemical mechanisms of HIV induced T cell apoptosis, Cell Death Differ., 8 (2001), 127–136. http://doi.org/10.1038/sj.cdd.4400822 doi: 10.1038/sj.cdd.4400822
|
| [21] |
R. L. Fan, Y. P. Dong, G. Huang, Y. Takeuchi, Apoptosis in virus infection dynamics models, J. Biol. Dynam., 8 (2014), 20–41. http://doi.org/10.1080/17513758.2014.895433 doi: 10.1080/17513758.2014.895433
|
| [22] |
M. Li, K. Guo, W. B. Ma, Uniform persistence and global attractivity in a delayed virus dynamic model with apoptosis and both virus-to-cell and cell-to-cell infections, Mathematics, 10 (2022), 975. http://doi.org/10.3390/math10060975 doi: 10.3390/math10060975
|
| [23] |
J. A. Zack, S. J. Arrigo, S. R. Weitsman, A. S. Go, A. Haislip, I. S. Y. Chen, HIV-1 entry into quiescent primary lymphocytes: molecular analysis reveals a labile, latent viral structure, Cell, 61 (1990), 213–222. http://doi.org/10.1016/0092-8674(90)90802-L doi: 10.1016/0092-8674(90)90802-L
|
| [24] |
J. A. Zack, A. M. Haislip, P. Krogstad, I. S. Chen, Incompletely reverse-transcribed human immunodeficiency virus type 1 genomes in quiescent cells can function as intermediates in the retroviral life cycle, J. Virol., 66 (1992), 1717–1725. http://doi.org/10.1128/jvi.66.3.1717-1725.1992 doi: 10.1128/jvi.66.3.1717-1725.1992
|
| [25] |
P. Essunger, A. S. Perelson, Modeling HIV infection of CD4+ T-cell subpopulations, J. Theor. Biol., 170 (1994), 367–391. http://doi.org/10.1006/jtbi.1994.1199 doi: 10.1006/jtbi.1994.1199
|
| [26] |
P. K. Srivastava, P. Chandra, Modeling the dynamics of HIV and CD4+ T cells during primary infection, Nonlinear Anal.-Real., 11 (2010), 612–618. http://doi.org/10.1016/j.nonrwa.2008.10.037 doi: 10.1016/j.nonrwa.2008.10.037
|
| [27] |
H. F. Xu, Q. X. Zhu, W. X. Zheng, Exponential stability of stochastic nonlinear delay systems subject to multiple periodic impulses, IEEE T. Automat. Contr., 69 (2024), 2621–2628. http://doi.org/10.1109/TAC.2023.3335005 doi: 10.1109/TAC.2023.3335005
|
| [28] |
L. C. Feng, Q. N. Zhang, C. Y. Zhang, D. X. Gong, Polynomial noises for nonlinear systems with nonlinear impulses and time-varying delays, Mathematics, 10 (2022), 1525. http://doi.org/10.3390/math10091525 doi: 10.3390/math10091525
|
| [29] |
Y. X. Zhao, L. S. Wang, Practical exponential stability of impulsive stochastic food chain system with time-varying delays, Mathematics, 11 (2023), 147. http://doi.org/10.3390/math11010147 doi: 10.3390/math11010147
|
| [30] | M. Martcheva, An introduction to mathematical epidemiology, New York: Springer, 2015. https://doi.org/10.1007/978-1-4899-7612-3 |
| [31] |
S. S. Chen, C. Y. Cheng, Y. Takeuchi, Stability analysis in delayed within-host viral dynamics with both viral and cellular infections, J. Math. Anal. Appl., 442 (2016), 642–672. http://doi.org/10.1016/j.jmaa.2016.05.003 doi: 10.1016/j.jmaa.2016.05.003
|
| [32] |
S. Pan, S. P. Chakrabarty, Threshold dynamics of HCV model with cell-to-cell transmission and a non-cytolytic cure in the presence of humoral immunity, Commun. Nonlinear Sci., 61 (2018), 180–197. http://doi.org/10.1016/j.cnsns.2018.02.010 doi: 10.1016/j.cnsns.2018.02.010
|
| [33] | J. K. Hale, S. M. V. Lunel, Introduction to functional differential equations, New York: Springer, 1993. https://doi.org/10.1007/978-1-4612-4342-7 |
| [34] |
H. R. Thieme, Convergence results and a Poincaré-Bendixson trichotomy for asymptotically autonomous differential equations, J. Math. Biol., 30 (1992), 755–763. http://doi.org/10.1007/BF00173267 doi: 10.1007/BF00173267
|
| [35] |
P. Wu, H. Y. Zhao, Mathematical analysis of an age-structured HIV/AIDS epidemic model with HAART and spatial diffusion, Nonlinear Anal.-Real., 60 (2021), 103289. http://doi.org/10.1016/j.nonrwa.2021.103289 doi: 10.1016/j.nonrwa.2021.103289
|
| [36] |
G. D. Li, Y. Zhang, Y. J. Guan, W. J. Li, Stability analysis of multi-point boundary conditions for fractional differential equation with non-instantaneous integral impulse, Math. Biosci. Eng., 20 (2023), 7020–7041. http://doi.org/10.3934/mbe.2023303 doi: 10.3934/mbe.2023303
|
| [37] |
T. T. Zheng, L. F. Nie, Z. D. Teng, Y. T. Luo, S. F. Wang, Analysis of an age-structured dengue model with multiple strains and cross immunity, Electron. J. Qual. Theory Differ. Equ., 2021 (2021), 50. http://doi.org/10.14232/ejqtde.2021.1.50 doi: 10.14232/ejqtde.2021.1.50
|
| [38] |
P. Wu, Z. S. Feng, X. B. Zhang, Global dynamics of an age-space structured HIV/AIDS model with viral load-dependent infection and conversion rates, J. Comput. Appl. Math., 412 (2022), 114309. http://doi.org/10.1016/j.cam.2022.114309 doi: 10.1016/j.cam.2022.114309
|
| [39] |
L. Q. Zhou, Z. X. Zhao, Q. X. Zhu, R. Zhou, T. W. Huang, Global polynomial stabilization of impulsive neural networks with bidirectional proportional delays, IEEE T. Netw. Sci. Eng., 11 (2024), 471–484. http://doi.org/10.1109/tnse.2023.3300907 doi: 10.1109/tnse.2023.3300907
|
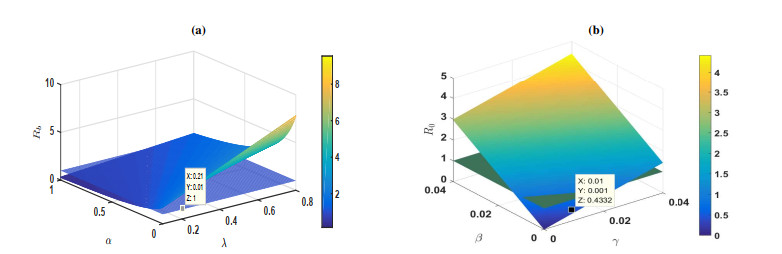
Figures(5) / Tables(1)

Ru Meng, Yantao Luo, Tingting Zheng. Stability analysis for a HIV model with cell-to-cell transmission, two immune responses and induced apoptosis[J]. AIMS Mathematics, 2024, 9(6): 14786-14806. doi: 10.3934/math.2024719







 DownLoad:
DownLoad: