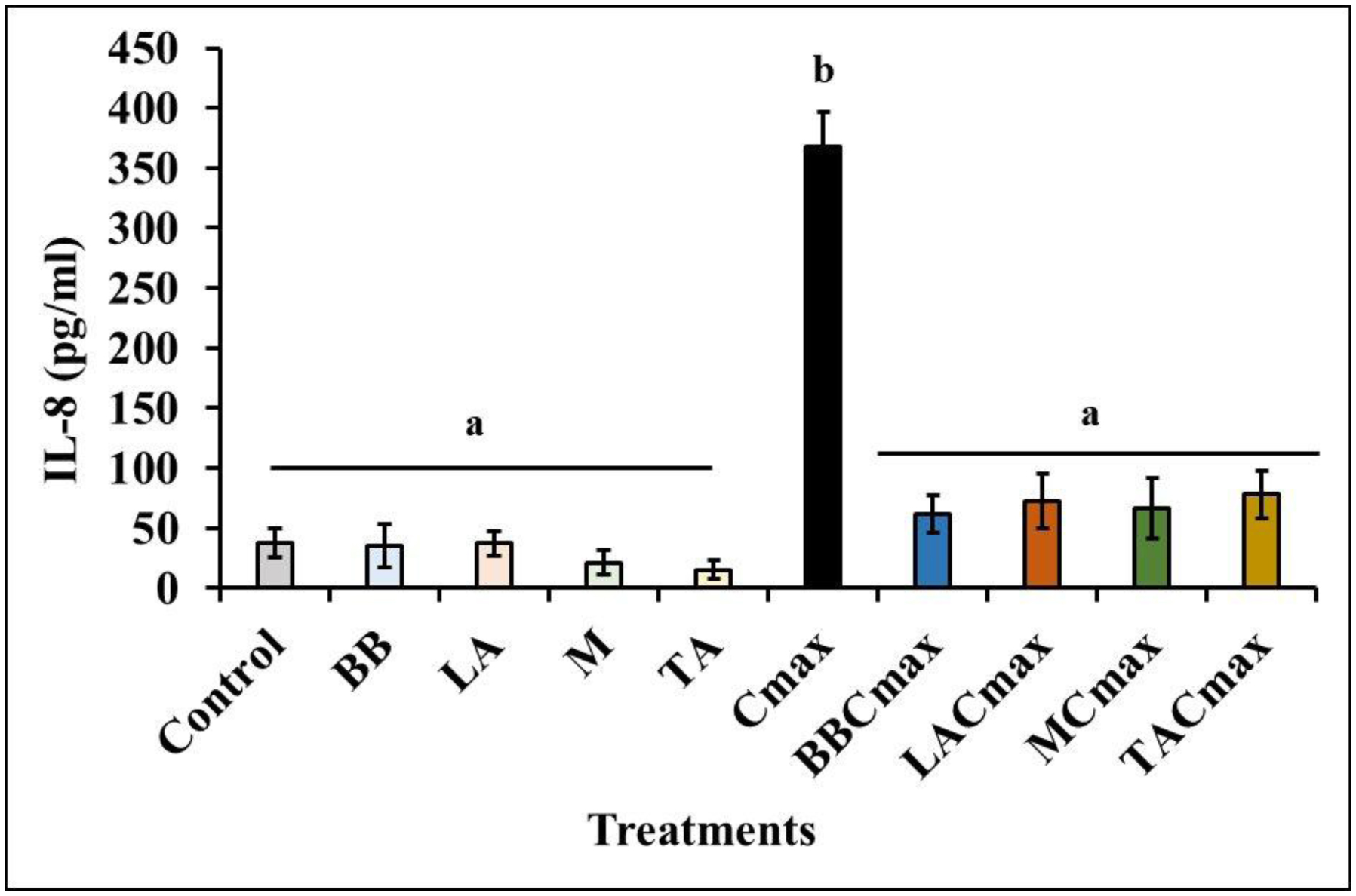
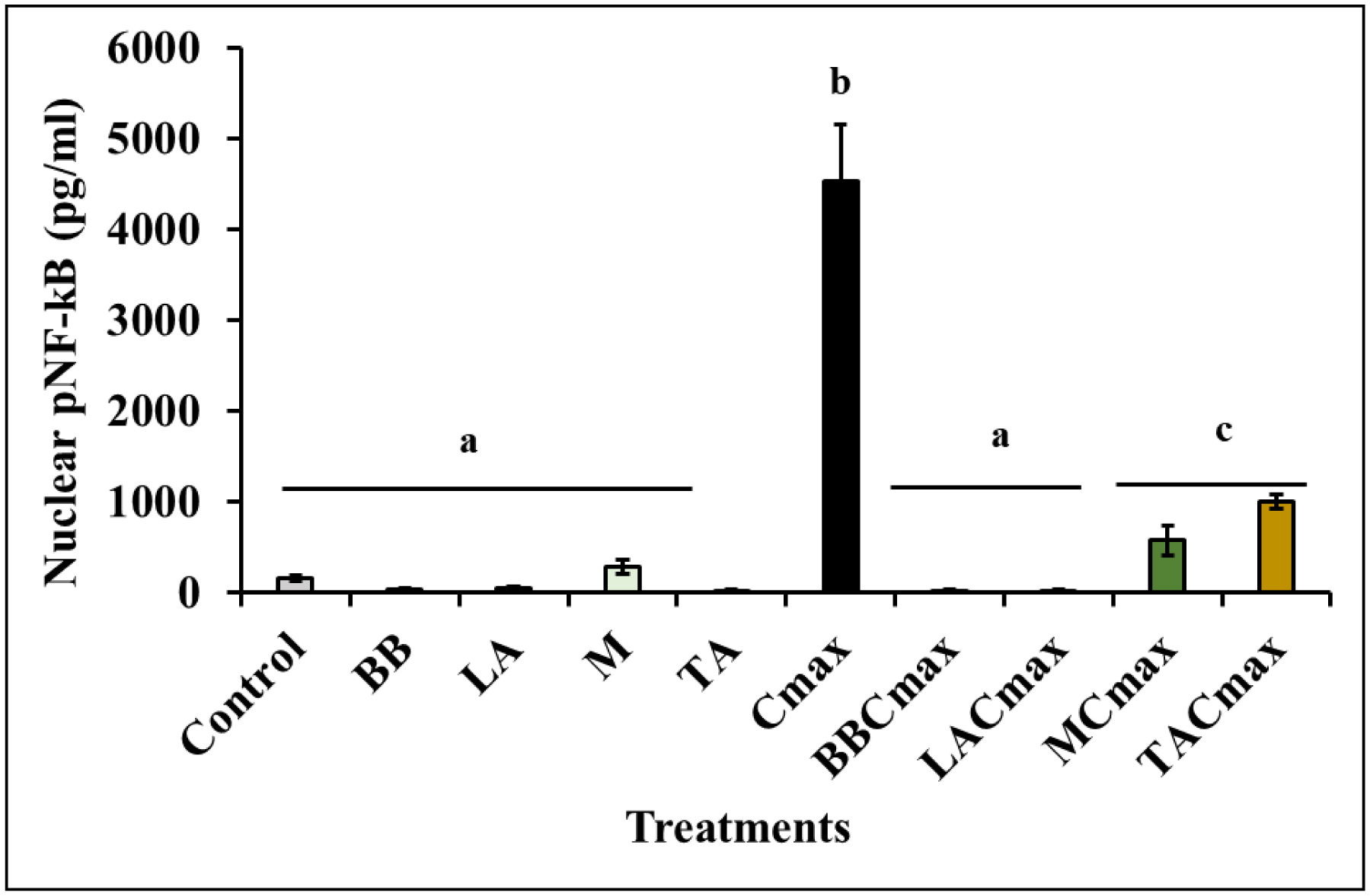
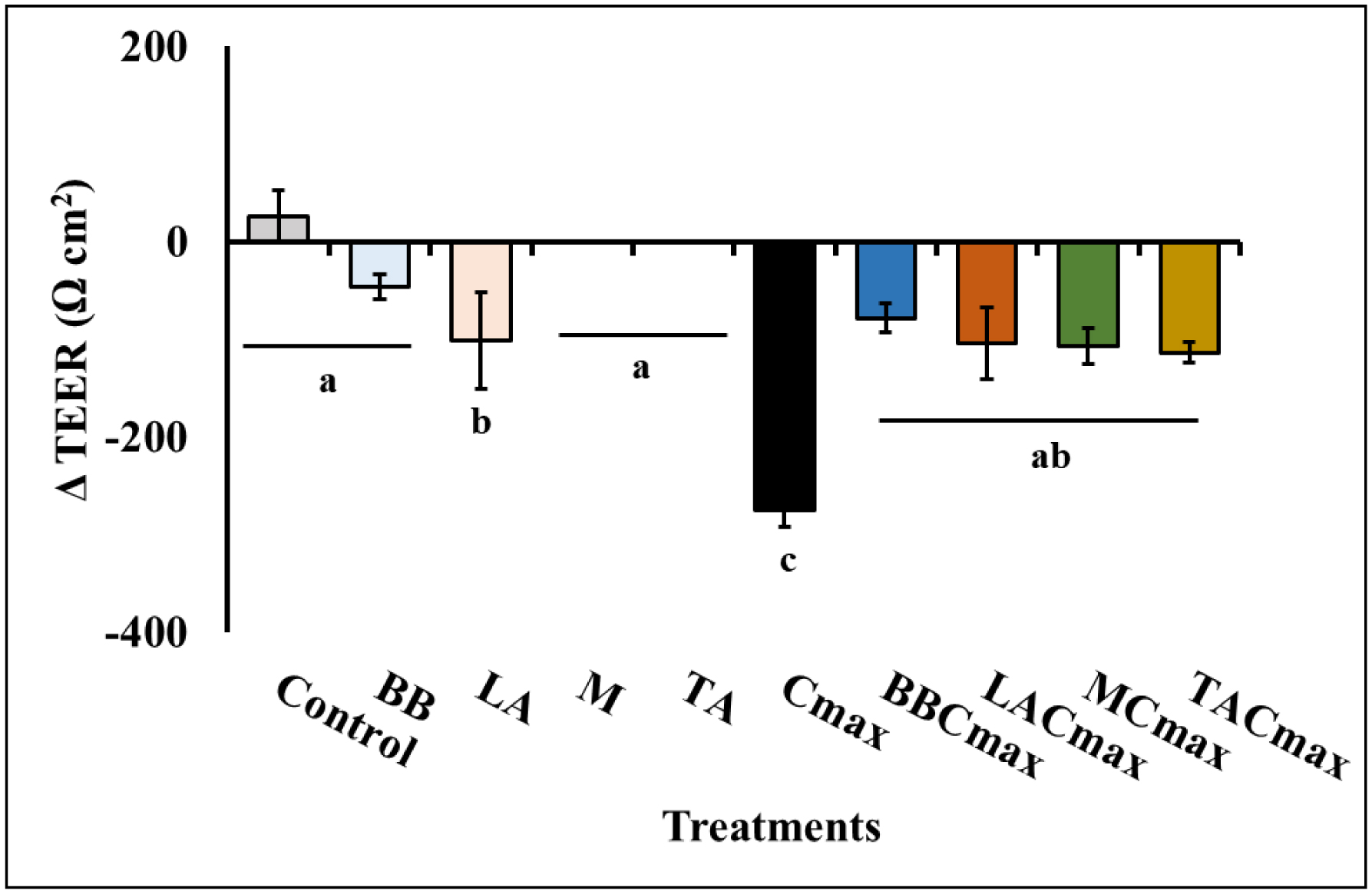
Chronic inflammation is identified to be an underlying pathophysiology in different conditions including inflammatory bowel disease (IBD). Since the aberrant interaction of the mucosal immune system with the dysbiotic flora has been reported to contribute to IBD development, probiotics have been studied for potential prophylaxis and treatment. In this regard, fermented dairy foods are a rich source of probiotics and bioactive compounds. However, limited studies have determined the impact of fermented dairy products in the context of chronic inflammation. In particular, a potential role for dairy starter cultures is not well studied. Hence, in this study we evaluated the anti-inflammatory effect of two cheese starter cultures (Lactococcus lactis subsp. lactis M58 and Streptococcus thermophilus TA 61) in comparison with commercial probiotic strains (Bifidobacterium animalis subsp. lactis BB-12, Lactobacillus acidophilus LA-5) using the Cmax-induced Caco-2 inflammation model. Specifically, we characterized their ability to attenuate inflammatory response via modulation of IL-8 secretion, NF-κB activation, barrier integrity (TEER), and tight junction gene expression. Overall, pre-exposure to the starter cultures before Cmax treatment significantly reduced the activation and nuclear translocation of NF-κB, compared to cytokine control (P < 0.05). Further, the reduction in pNF-κB was found to be associated with a significant reduction in IL 8 secretion (P < 0.05). Moreover, the cultures protected the Caco-2 monolayer from inflammation-induced increase in permeability by upregulating the genes associated with ZO-1 and occludin production. Furthermore, the protective effect of the starter cultures was comparable to that of the commercial probiotics with known anti-inflammatory properties. Therefore, cheese starter cultures could be a potential strategy against chronic gut inflammation.
Citation: Deepa Kuttappan, Sulthana Humayoon Muttathukonam, Mary Anne Amalaradjou. Cheese starter cultures attenuate inflammation in the in vitro Caco-2 model[J]. AIMS Microbiology, 2025, 11(2): 369-387. doi: 10.3934/microbiol.2025017
Chronic inflammation is identified to be an underlying pathophysiology in different conditions including inflammatory bowel disease (IBD). Since the aberrant interaction of the mucosal immune system with the dysbiotic flora has been reported to contribute to IBD development, probiotics have been studied for potential prophylaxis and treatment. In this regard, fermented dairy foods are a rich source of probiotics and bioactive compounds. However, limited studies have determined the impact of fermented dairy products in the context of chronic inflammation. In particular, a potential role for dairy starter cultures is not well studied. Hence, in this study we evaluated the anti-inflammatory effect of two cheese starter cultures (Lactococcus lactis subsp. lactis M58 and Streptococcus thermophilus TA 61) in comparison with commercial probiotic strains (Bifidobacterium animalis subsp. lactis BB-12, Lactobacillus acidophilus LA-5) using the Cmax-induced Caco-2 inflammation model. Specifically, we characterized their ability to attenuate inflammatory response via modulation of IL-8 secretion, NF-κB activation, barrier integrity (TEER), and tight junction gene expression. Overall, pre-exposure to the starter cultures before Cmax treatment significantly reduced the activation and nuclear translocation of NF-κB, compared to cytokine control (P < 0.05). Further, the reduction in pNF-κB was found to be associated with a significant reduction in IL 8 secretion (P < 0.05). Moreover, the cultures protected the Caco-2 monolayer from inflammation-induced increase in permeability by upregulating the genes associated with ZO-1 and occludin production. Furthermore, the protective effect of the starter cultures was comparable to that of the commercial probiotics with known anti-inflammatory properties. Therefore, cheese starter cultures could be a potential strategy against chronic gut inflammation.

| [1] |
Netea MG, Balkwill F, Chonchol, et al. (2017) A guiding map for inflammation. Nat Immunol 18: 826-831. https://doi.org/10.1038/ni.3790 
|
| [2] |
Agrawal M, Allin KH, Petralia F, et al. (2022) Multiomics to elucidate inflammatory bowel disease risk factors and pathways. Nat Rev Gastroenterol Hepatol 19: 399-409. https://doi.org/10.1038/s41575-022-00593-y
|
| [3] |
Furman D, Campisi J, Verdin E, et al. (2019) Chronic inflammation in the etiology of disease across the life span. Nat Med 25: 1822-1832. https://doi.org/10.1038/s41591-019-0675-0
|
| [4] |
Massimino L, Lamparelli LA, Houshyar Y, et al. (2021) The inflammatory bowel disease transcriptome and metatranscriptome meta-analysis (IBD TaMMA) framework. Nat Comput Sci 1: 511-515. https://doi.org/10.1038/s43588-021-00114-y
|
| [5] |
McCole DF (2014) IBD candidate genes and intestinal barrier regulation. Inflamm Bowel Dis 20: 1829-1849. https://doi.org/10.1097/MIB.0000000000000090
|
| [6] |
Ramos GP, Papadakis KA (2019) Mechanisms of disease: inflammatory bowel diseases. Mayo Clin Proceedings 94: 155-165. https://doi.org/10.1016/j.mayocp.2018.09.013
|
| [7] |
Vancamelbeke M, Vanuytsel T, Farré R, et al. (2017) Genetic and transcriptomic bases of intestinal epithelial barrier dysfunction in inflammatory bowel disease. Inflamm Bowel Dis 23: 1718-1729. https://doi.org/10.1097/MIB.0000000000001246
|
| [8] |
Van De Walle J, Hendrickx A, Romier B, et al. (2010) Inflammatory parameters in Caco-2 cells: Effect of stimuli nature, concentration, combination, and cell differentiation. In Vitro Toxicol 24: 1441-1449. https://doi.org/10.1016/j.tiv.2010.04.002
|
| [9] |
Danese S (2008) Nonimmune cells in inflammatory bowel disease: from victim to villain. Trends Immunol 29: 555-564. http://doi.org/10.1016/j.it.2008.07.009
|
| [10] |
Allaire JM, Morampudi V, Crowley SM, et al. (2018) Frontline defenders: goblet cell mediators dictate host-microbe interactions in the intestinal tract during health and disease. Am J Physiol Gastrointest Liver Physiol 314: G360-G377. https://doi.org/10.1152/ajpgi.00181.2017
|
| [11] |
Koch S, Nusrat A, Parkos CA (2013) The epithelial barrier. Molecular Genetics of Inflammatory Bowel Disease . New York, NY: Springer New York 265-280. https://doi.org/10.1007/978-1-4614-8256-7_13
|
| [12] |
Wallace KL, Zheng LB, Kanazawa Y, et al. (2014) Immunopathology of inflammatory bowel disease. WJG 20: 6. https://doi.org/10.3748/wjg.v20.i1.6
|
| [13] |
Barbara G, Barbaro MR, Fuschi D, et al. (2021) Inflammatory and microbiota-related regulation of the intestinal epithelial barrier. Front Nutr 8: 718356. https://doi.org/10.3389/fnut.2021.718356
|
| [14] |
Mahapatro M, Erkert L, Becker C (2021) Cytokine-mediated crosstalk between immune cells and epithelial cells in the gut. Cells 10: 111. https://doi.org/10.3390/cells10010111
|
| [15] |
Yao Y, Shang W, Bao L, et al. (2024) Epithelial-immune cell crosstalk for intestinal barrier homeostasis. Eur J Immunol 54: 2350631. https://doi.org/10.1002/eji.202350631
|
| [16] |
Akira S, Uematsu S, Takeuchi O (2006) Pathogen recognition and innate immunity. Cell 124: 783-801. https://doi.org/10.1016/j.cell.2006.02.015
|
| [17] |
Nakase H, Sato N, Mizuno N, et al. (2022) The influence of cytokines on the complex pathology of ulcerative colitis. Autoimmun Rev 21: 103017. https://doi.org/10.1016/j.autrev.2021.103017
|
| [18] |
Leppkes M, Neurath MF (2020) Cytokines in inflammatory bowel diseases–update 2020. Pharmacol Res 158: 104835. https://doi.org/10.1016/j.phrs.2020.104835
|
| [19] |
Ma Y, Yang D, Huang J, et al. (2024) Probiotics for inflammatory bowel disease: Is there sufficient evidence?. Open Life Sci 19: 20220821. https://doi.org/10.1515/biol-2022-0821
|
| [20] |
Chae JM, Heo W, Cho HT, et al. (2018) Effects of orally-administered Bifidobacterium animalis subsp. lactis strain BB12 on dextran sodium sulfate-induced colitis in mice. J Microbiol Biotechnol 28: 1800-1805. https://doi.org/10.4014/jmb.1805.05072
|
| [21] | Wildt S, Nordgaard I, Hansen U, et al. (2011) A randomised double-blind placebo-controlled trial with Lactobacillus acidophilus La-5 and Bifidobacterium animalis subsp. lactis BB-12 for maintenance of remission in ulcerative colitis. JCC 5: 115-121. https://doi.org/10.1093/ecco-jcc/jjac104 |
| [22] | Feathers A, Grigoryan Z, Falzon L, et al. (2023) S1004 The role of fermented food in inflammatory bowel disease treatment: A systematic review of randomized trials. ACG 118: S760-S761. https://doi.org/10.14309/01.ajg.0000953656.43074.2e |
| [23] |
Illikoud N, Mantel M, Rolli-Derkinderen M, et al. (2022) Dairy starters and fermented dairy products modulate gut mucosal immunity. Immunol Lett 251: 91-102. https://doi.org/10.1016/j.imlet.2022.11.002
|
| [24] |
García-Burgos M, Moreno-Fernández J, Alférez MJ, et al. (2020) New perspectives in fermented dairy products and their health relevance. J Funct Foods 72: 104059. https://doi.org/10.1016/j.jff.2020.104059
|
| [25] | Moreno Y, Collado MC, Ferrús MA, et al. (2006) Viability assessment of lactic acid bacteria in commercial dairy products stored at 4 C using LIVE/DEAD® BacLightTM staining and conventional plate counts. IJFST 41: 275-280. https://doi.org/10.1111/j.1365-2621.2005.01060.x |
| [26] |
Moser A, Schafroth K, Meile L, et al. (2018) Population dynamics of Lactobacillus helveticus in Swiss Gruyère-type cheese manufactured with natural whey cultures. Front Microbiol 9: 637. https://doi.org/10.3389/fmicb.2018.00637
|
| [27] | De Freitas I, Pinon N, Thierry A, et al. (2007) In depth dynamic characterization of French PDO Cantal cheese made from raw milk. Diary Sci Technol 87: 97-117. https://doi.org/10.1051/lait:2007007 |
| [28] |
Poveda JM, Sousa MJ, Cabezas L, et al. (2003) Preliminary observations on proteolysis in Manchego cheese made with a defined-strain starter culture and adjunct starter (Lactobacillus plantarum) or a commercial starter. Int Dairy J 13: 169-178. https://doi.org/10.1016/S0958-6946(02)00150-4
|
| [29] | Ito M, Ohishi K, Yoshida Y, et al. (2008) Preventive effect of Streptococcus thermophilus YIT 2001 on dextran sulfate sodium-induced colitis in mice. BBBIEJ 72: 2543-2547. https://doi.org/10.1271/bbb.80240 |
| [30] |
Ogita T, Nakashima M, Morita H, et al. (2011) Streptococcus thermophilus ST28 ameliorates colitis in mice partially by suppression of inflammatory Th17 cells. Biomed Res Int 1: 378417. https://doi.org/10.1155/2011/378417
|
| [31] |
Berlec A, Perše M, Ravnikar M, et al. (2017) Dextran sulphate sodium colitis in C57BL/6J mice is alleviated by Lactococcus lactis and worsened by the neutralization of tumor necrosis factor α. Int Immunopharmacol 43: 219-226. https://doi.org/10.1016/j.intimp.2016.12.027
|
| [32] |
Ambalam P, Kondepudi KK, Balusupati P, et al. (2015) Prebiotic preferences of human lactobacilli strains in co-culture with bifidobacteria and antimicrobial activity against Clostridium difficile. J Appl Microbiol 119: 1672-1682. https://doi.org/10.1111/jam.12953
|
| [33] |
de Carvalho Lima KG, Kruger MF, Behrens J, et al. (2009) Evaluation of culture media for enumeration of Lactobacillus acidophilus, Lactobacillus casei and Bifidobacterium animalis in the presence of Lactobacillus delbrueckii subsp bulgaricus and Streptococcus thermophilus. LWT-Food Sci Technol 42: 491-495. https://doi.org/10.1016/j.lwt.2008.08.011
|
| [34] |
Fachin L, Moryia J, Gândara ALN, et al. (2008) Evaluation of culture media for counts of Bifidobacterium animalis subsp. lactis Bb 12in yoghurt after refrigerated storage. Braz J Microbiol 39: 357-361. https://doi.org/10.1590/S1517-83822008000200029
|
| [35] |
Putt KK, Pei R, White HM, et al. (2017) Yogurt inhibits intestinal barrier dysfunction in Caco-2 cells by increasing tight junctions. Food Funct 8: 406-414. https://doi.org/10.1039/C6FO01592A
|
| [36] |
Ciorba MA, Riehl TE, Rao MS, et al. (2012) Lactobacillus probiotic protects intestinal epithelium from radiation injury in a TLR-2/cyclo-oxygenase-2-dependent manner. Gut 61: 829-838. https://doi.org/10.1136/gutjnl-2011-300367
|
| [37] |
Dimitrov Z, Gotova I, Chorbadjiyska E (2014) In vitro characterization of the adhesive factors of selected probiotics to Caco-2 epithelium cell line. Biotechnol Biotechnol Equip 28: 1079-1083. https://doi.org/10.1080/13102818.2014.969948
|
| [38] |
Buagaew A, Poomipark N (2020) Protective effect of piperine from Piper chaba fruits on LPS-induced inflammation in human intestinal cell line. J Med Plants Res 14: 438-444. https://doi.org/10.5897/JMPR2020.6996
|
| [39] |
Simmons LJ, Surles-Zeigler MC, Li Y, et al. (2016) Regulation of inflammatory responses by neuregulin-1 in brain ischemia and microglial cells in vitro involves the NF-kappa B pathway. J Neuroinflammation 13: 1-15. https://doi.org/10.1186/s12974-016-0703-7
|
| [40] |
Van K, Burns JL, Monk JM (2024) Effect of short-chain fatty acids on inflammatory and metabolic function in an obese skeletal muscle cell culture model. Nutrients 16: 500. https://doi.org/10.3390/nu16040500
|
| [41] | Matsumoto S, Hara T, Hori T, et al. (2005) Probiotic Lactobacillus-induced improvement in murine chronic inflammatory bowel disease is associated with the down-regulation of pro-inflammatory cytokines in lamina propria mononuclear cells. CEI 140: 417-426. https://doi.org/10.1111/j.1365-2249.2005.02790.x |
| [42] |
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 25: 402-408. https://doi.org/10.1006/meth.2001.1262
|
| [43] |
Ferruzza S, Rossi C, Scarino ML, et al. (2012) A protocol for differentiation of human intestinal Caco-2 cells in asymmetric serum-containing medium. In Vitro Toxicol 26: 1252-1255. https://doi.org/10.1016/j.tiv.2012.01.008
|
| [44] | Lea T (2015) Caco-2 cell line. The Impact of Food Bioactives on Health: in vitro and ex vivo models . Springer 103-111. https://doi.org/10.1007/978-3-319-16104-4 |
| [45] | Guan Q, Zhang J (2017) Recent advances: the imbalance of cytokines in the pathogenesis of inflammatory bowel disease. Mediat Inflamm 1: 4810258. https://doi.org/10.1155/2017/4810258 |
| [46] |
Xu P, Elamin E, Elizalde M, et al. (2019) Modulation of intestinal epithelial permeability by plasma from patients with Crohn's disease in a three-dimensional cell culture model. Sci Rep 9: 2030. https://doi.org/10.1038/s41598-018-38322-8
|
| [47] |
Aggeletopoulou I, Kalafateli M, Tsounis EP, et al. (2024) Exploring the role of IL-1β in inflammatory bowel disease pathogenesis. Front Med 11: 1307394. https://doi.org/10.3389/fmed.2024.1307394
|
| [48] |
Kurumi H, Yokoyama Y, Hirano T, et al. (2024) Cytokine profile in predicting the effectiveness of advanced therapy for ulcerative colitis: A narrative review. Biomedicines 12: 952. https://doi.org/10.3390/biomedicines12050952
|
| [49] |
Kaminsky LW, Al-Sadi R, Ma TY (2021) IL-1β and the intestinal epithelial tight junction barrier. Front Immunol 12: 767456. https://doi.org/10.3389/fimmu.2021.767456
|
| [50] |
Meyer F, Wendling D, Demougeot C, et al. (2023) Cytokines and intestinal epithelial permeability: A systematic review. Autoimmun Rev 22: 103331. https://doi.org/10.1016/j.autrev.2023.103331
|
| [51] |
Subramanian S, Rhodes JM, Hart AC, et al. (2008) Characterization of epithelial IL-8 response to inflammatory bowel disease mucosal E. coli and its inhibition by mesalamine. Inflamm Bowel Dis 14: 162-175. https://doi.org/10.1002/ibd.20296
|
| [52] |
Hoffmann E, Dittrich-Breiholz O, Holtmann H, et al. (2002) Multiple control of interleukin-8 gene expression. J Leukoc Biol 72: 847-855. https://doi.org/10.1189/jlb.72.5.847
|
| [53] |
Zhu Y, Yang S, Zhao N, et al. (2021) CXCL8 chemokine in ulcerative colitis. Biomed Pharmacother 138: 111427. https://doi.org/10.1016/j.biopha.2021.111427
|
| [54] | Yoshida N, Katada K, Handa O, et al. (2007) Interleukin-8 production via protease-activated receptor 2 in human esophageal epithelial cells. Int J Mol Med 19: 335-340. https://doi.org/10.3892/ijmm.19.2.335 |
| [55] | Bie Y, Ge W, Yang Z, et al. (2019) The crucial role of CXCL8 and its receptors in colorectal liver metastasis. Dis Markers 1: 8023460. https://doi.org/10.1155/2019/8023460 |
| [56] |
Wanninger J, Neumeier M, Weigert J, et al. (2009) Adiponectin-stimulated CXCL8 release in primary human hepatocytes is regulated by ERK1/ERK2, p38 MAPK, NF-κB, and STAT3 signaling pathways. Am J Physiol Gastrointest Liver Physiol 297: G611-G618. https://doi.org/10.1152/ajpgi.90644.2008
|
| [57] |
Brat DJ, Bellail AC, Van Meir EG (2005) The role of interleukin-8 and its receptors in gliomagenesis and tumoral angiogenesis. Neuro-oncology 7: 122-133. https://doi.org/10.1215/S1152851704001061
|
| [58] |
Ha H, Debnath B, Neamati N (2017) Role of the CXCL8-CXCR1/2 axis in cancer and inflammatory diseases. Theranostics 7: 1543. https://doi.org/10.7150/thno.15625
|
| [59] |
Waugh DJ, Wilson C (2008) The interleukin-8 pathway in cancer. Clin Cancer Res 14: 6735-6741. https://doi.org/10.1158/1078-0432.CCR-07-4843
|
| [60] |
Russo RC, Garcia CC, Teixeira MM, et al. (2014) The CXCL8/IL-8 chemokine family and its receptors in inflammatory diseases. Expert Rev Clin Immunol 10: 593-619. https://doi.org/10.1586/1744666X.2014.894886
|
| [61] |
Ibusuki K, Sakiyama T, Kanmura S, et al. (2015) Human neutrophil peptides induce interleukin-8 in intestinal epithelial cells through the P2 receptor and ERK1/2 signaling pathways. Int J Mol Med 35: 1603-1609. https://doi.org/10.3892/ijmm.2015.2156
|
| [62] | Bruno ME, Rogier EW, Arsenescu RI, et al. (2015) Correlation of biomarker expression in colonic mucosa with disease phenotype in Crohn's disease and ulcerative colitis. DDS 60: 2976-2984. https://doi.org/10.1007/s10620-015-3700-2 |
| [63] |
Zahn A, Giese T, Karner M, et al. (2009) Transcript levels of different cytokines and chemokines correlate with clinical and endoscopic activity in ulcerative colitis. BMC Gastroenterol 9: 1-7. https://doi.org/10.1186/1471-230X-9-13
|
| [64] |
Cai Z, Wang S, Li J (2021) Treatment of inflammatory bowel disease: a comprehensive review. Front Med 8: 765474. https://doi.org/10.3389/fmed.2021.765474
|
| [65] |
Shin JY, Wey M, Umutesi HG, et al. (2016) Thiopurine prodrugs mediate immunosuppressive effects by interfering with Rac1 protein function. JBC 291: 13699-13714. https://doi.org/10.1074/jbc.M115.694422
|
| [66] |
Van Dieren JM, Kuipers EJ, Samsom JN, et al. (2006) Revisiting the immunomodulators tacrolimus, methotrexate, and mycophenolate mofetil: their mechanisms of action and role in the treatment of IBD. Inflamm Bowel Dis 12: 311-327. https://doi.org/10.1097/01.MIB.0000209787.19952.53
|
| [67] |
Mardini HE, Grigorian AY (2014) Probiotic mix VSL# 3 is effective adjunctive therapy for mild to moderately active ulcerative colitis: a meta-analysis. Inflamm Bowel Dis 20: 1562-1567. https://doi.org/10.1097/MIB.0000000000000084
|
| [68] |
Zhang XF, Guan XX, Tang YJ, et al. (2021) Clinical effects and gut microbiota changes of using probiotics, prebiotics or synbiotics in inflammatory bowel disease: A systematic review and meta-analysis. Eur J Nutr 60: 2855-2875. https://doi.org/10.1007/s00394-021-02503-5
|
| [69] |
Ganji-Arjenaki M, Rafieian-Kopaei M (2018) Probiotics are a good choice in remission of inflammatory bowel diseases: A meta analysis and systematic review. J Cell Physiol 233: 2091-2103. https://doi.org/10.1002/jcp.25911
|
| [70] |
Yılmaz İ, Dolar ME, Özpınar H (2018) Effect of administering kefir on the changes in fecal microbiota and symptoms of inflammatory bowel disease: A randomized controlled trial. Turk J Gastroenterol 30: 242. https://doi.org/10.5152/tjg.2018.18227
|
| [71] |
Pápai G, Torres-Maravilla E, Chain F, et al. (2021) The administration matrix modifies the beneficial properties of a probiotic mix of Bifidobacterium animalis subsp. lactis BB-12 and Lactobacillus acidophilus LA-5. Probiotics Antimicrob Proteins 13: 484-494. https://doi.org/10.1007/s12602-020-09702-2
|
| [72] |
Splichal I, Donovan SM, Kindlova Z, et al. (2023) Release of HMGB1 and toll-like receptors 2, 4, and 9 signaling are modulated by Bifidobacterium animalis subsp. lactis BB-12 and Salmonella Typhimurium in a gnotobiotic piglet model of preterm infants. Int J Mol Sci 24: 2329. https://doi.org/10.3390/ijms24032329
|
| [73] |
Luerce TD, Gomes-Santos AC, Rocha CS, et al. (2014) Anti-inflammatory effects of Lactococcus lactis NCDO 2118 during the remission period of chemically induced colitis. Gut Pathog 6: 1-11. https://doi.org/10.1186/1757-4749-6-33
|
| [74] |
Kawahara M, Nemoto M, Nakata T, et al. (2015) Anti-inflammatory properties of fermented soy milk with Lactococcus lactis subsp. lactis S-SU2 in murine macrophage RAW264. 7 cells and DSS-induced IBD model mice. Int Immunopharmacol 26: 295-303. https://doi.org/10.1016/j.intimp.2015.04.004
|
| [75] | Gotova I, Dimitrov Z, Najdenski H (2017) Selected Lactobacillus bulgaricus and Streptococcus thermophilus strains from bulgarian yogurt demonstrate significant anti-inflammatory potential. Acta Microbiol Bulg 33: 7. |
| [76] |
Karin M, Greten FR (2005) NF-κB: linking inflammation and immunity to cancer development and progression. Nat Rev Immunol 5: 749-759. https://doi.org/10.1038/nri1703
|
| [77] |
Taniguchi K, Karin M (2018) NF-κB, inflammation, immunity, and cancer: coming of age. Nat Rev Immunol 18: 309-324. https://doi.org/10.1038/nri.2017.142
|
| [78] |
Hayden MS, Ghosh S (2008) Shared principles in NF-κB signaling. Cell 132: 344-362. https://doi.org/10.1016/j.cell.2008.01.020
|
| [79] |
McDaniel DK, Eden K, Ringel VM, et al. (2016) Emerging roles for noncanonical NF-κB signaling in the modulation of inflammatory bowel disease pathobiology. Inflamm Bowel Dis 22: 2265-2279. https://doi.org/10.1097/MIB.0000000000000858
|
| [80] |
Heuvelin E, Lebreton C, Grangett, C, et al. (2009) Mechanisms involved in alleviation of intestinal inflammation by Bifidobacterium breve soluble factors. PLoS One 4: e5184. https://doi.org/10.1371/journal.pone.0005184
|
| [81] |
Meng H, Ba Z, Lee Y, et al. (2017) Consumption of Bifidobacterium animalis subsp. lactis BB-12 in yogurt reduced expression of TLR-2 on peripheral blood-derived monocytes and pro-inflammatory cytokine secretion in young adults. Eur J Nutr 56: 649-661. https://doi.org/10.1007/s00394-015-1109-5
|
| [82] |
Okada Y, Tsuzuki Y, Hokari R, et al. (2009) Anti-inflammatory effects of the genus Bifidobacterium on macrophages by modification of phospho-IκB and SOCS gene expression. Int J Exp Pathol 90: 131-140. https://doi.org/10.1111/j.1365-2613.2008.00632.x
|
| [83] |
Riedel CU, Foata F, Philippe D, et al. (2006) Anti-inflammatory effects of bifidobacteria by inhibition of LPS-induced NF-κB activation. WJG 12: 3729. https://doi.org/10.3748/wjg.v12.i23.3729
|
| [84] |
Tien MT, Girardin SE, Regnault B, et al. (2006) Anti-inflammatory effect of Lactobacillus casei on Shigella-infected human intestinal epithelial cells. J Immunol 176: 1228-1237. https://doi.org/10.4049/jimmunol.176.2.1228
|
| [85] | Barbara G (2006) Mucosal barrier defects in irritable bowel syndrome. Who left the door open?. ACG 101: 1295-1298. https://journals.lww.com/ajg/fulltext/2006/06000/Mucosal_Barrier_Defects_in_Irritable_Bowel.23.aspx |
| [86] |
Michielan A, D'Incà R (2015) Intestinal permeability in inflammatory bowel disease: pathogenesis, clinical evaluation, and therapy of leaky gut. Mediat Inflamm 1: 628157. https://doi.org/10.1155/2015/628157
|
| [87] |
Wang W, Chen L, Zhou R, et al. (2014) Increased proportions of Bifidobacterium and the Lactobacillus group and loss of butyrate-producing bacteria in inflammatory bowel disease. J Clin Microbiol 52: 398-406. https://doi.org/10.1128/jcm.01500-13
|
| [88] |
Zakostelska Z, Kverka M, Klimesova K, et al. (2011) Lysate of probiotic Lactobacillus casei DN-114 001 ameliorates colitis by strengthening the gut barrier function and changing the gut microenvironment. PloS One 6: e27961. https://doi.org/10.1371/journal.pone.0027961
|
| [89] |
Wang H, Fan C, Zhao Z, et al. (2022) Anti-inflammatory effect of Bifidobacterium animalis subsp. lactis A6 on DSS-induced colitis in mice. J App Microbiol 133: 2063-2073. https://doi.org/10.1111/jam.15681
|
| [90] |
Bailey JR, Vince V, Williams NA, et al. (2017) Streptococcus thermophilus NCIMB 41856 ameliorates signs of colitis in an animal model of inflammatory bowel disease. Benef Microbes 8: 605-614. https://doi.org/10.3920/BM2016.0110
|
| [91] | Anderson JM, Van Itallie CM (2009) Physiology and function of the tight junction. CSH Perspect Biol 1: a002584. |
| [92] |
Furuse M, Hirase T, Itoh M, et al. (1993) Occludin: a novel integral membrane protein localizing at tight junctions. JCB 123: 1777-1788. https://doi.org/10.1083/jcb.123.6.1777
|
| [93] |
Ikenouchi J, Umeda K, Tsukita S, et al. (2007) Requirement of ZO-1 for the formation of belt-like adherens junctions during epithelial cell polarization. JCB 176: 779-786. https://doi.org/10.1083/jcb.200612080
|
| [94] |
Wong V (1997) Phosphorylation of occludin correlates with occludin localization and function at the tight junction. Am J Physiol Cell Physiol 273: C1859-C1867. https://doi.org/10.1152/ajpcell.1997.273.6.C1859
|
| [95] |
Fanning AS, Ma TY, Anderson JM (2002) Isolation and functional characterization of the actin-binding region in the tight junction protein ZO-1. FASEB J 16: 1-23. https://doi.org/10.1096/fj.02-0121fje
|
| [96] |
Martínez C, Vicario M, Ramos L, et al. (2012) The jejunum of diarrhea-predominant irritable bowel syndrome shows molecular alterations in the tight junction signaling pathway that are associated with mucosal pathobiology and clinical manifestations. Am J Gastroenterol 107: 736-746. https://doi.org/10.1038/ajg.2011.472
|
| [97] |
Wilcz-Villega E, McClean S, O'sullivan M (2014) Reduced E-cadherin expression is associated with abdominal pain and symptom duration in a study of alternating and diarrhea predominant IBS. Neurogastroenterol Motil 26: 316-325. https://doi.org/10.1111/nmo.12262
|
| [98] |
Drago S, El Asmar R, Di Pierro M, et al. (2006) Gliadin, zonulin and gut permeability: Effects on celiac and non-celiac intestinal mucosa and intestinal cell lines. Scand J Gastroenterol 41: 408-419. https://doi.org/10.1080/00365520500235334
|
| [99] |
Anderson RC, Cookson AL, McNabb WC, et al. (2010) Lactobacillus plantarum MB452 enhances the function of the intestinal barrier by increasing the expression levels of genes involved in tight junction formation. BMC Microbiol 10: 1-11. https://doi.org/10.1186/1471-2180-10-316
|
| [100] |
Zhao L, Xie Q, Evivie SE, et al. (2021) Bifidobacterium dentium N8 with potential probiotic characteristics prevents LPS-induced intestinal barrier injury by alleviating the inflammatory response and regulating the tight junction in Caco-2 cell monolayers. Food Funct 12: 7171-7184. https://doi.org/10.1039/D1FO01164B
|
| [101] |
Do Carmo FLR, Rabah H, Cordeiro BF, et al. (2019) Probiotic Propionibacterium freudenreichii requires SlpB protein to mitigate mucositis induced by chemotherapy. Oncotarget 10: 7198. https://doi.org/10.18632/oncotarget.27319
|
| [102] |
Chen Y, Bi S, Zhang X, et al. (2025) Engineered probiotics remodel the intestinal epithelial barrier and enhance bacteriotherapy for inflammatory bowel diseases. Acta Biomaterialia 198: 467-481. https://doi.org/10.1016/j.actbio.2025.04.016
|
Figures(3) / Tables(2)

Deepa Kuttappan, Sulthana Humayoon Muttathukonam, Mary Anne Amalaradjou. Cheese starter cultures attenuate inflammation in the in vitro Caco-2 model[J]. AIMS Microbiology, 2025, 11(2): 369-387. doi: 10.3934/microbiol.2025017







 DownLoad:
DownLoad: