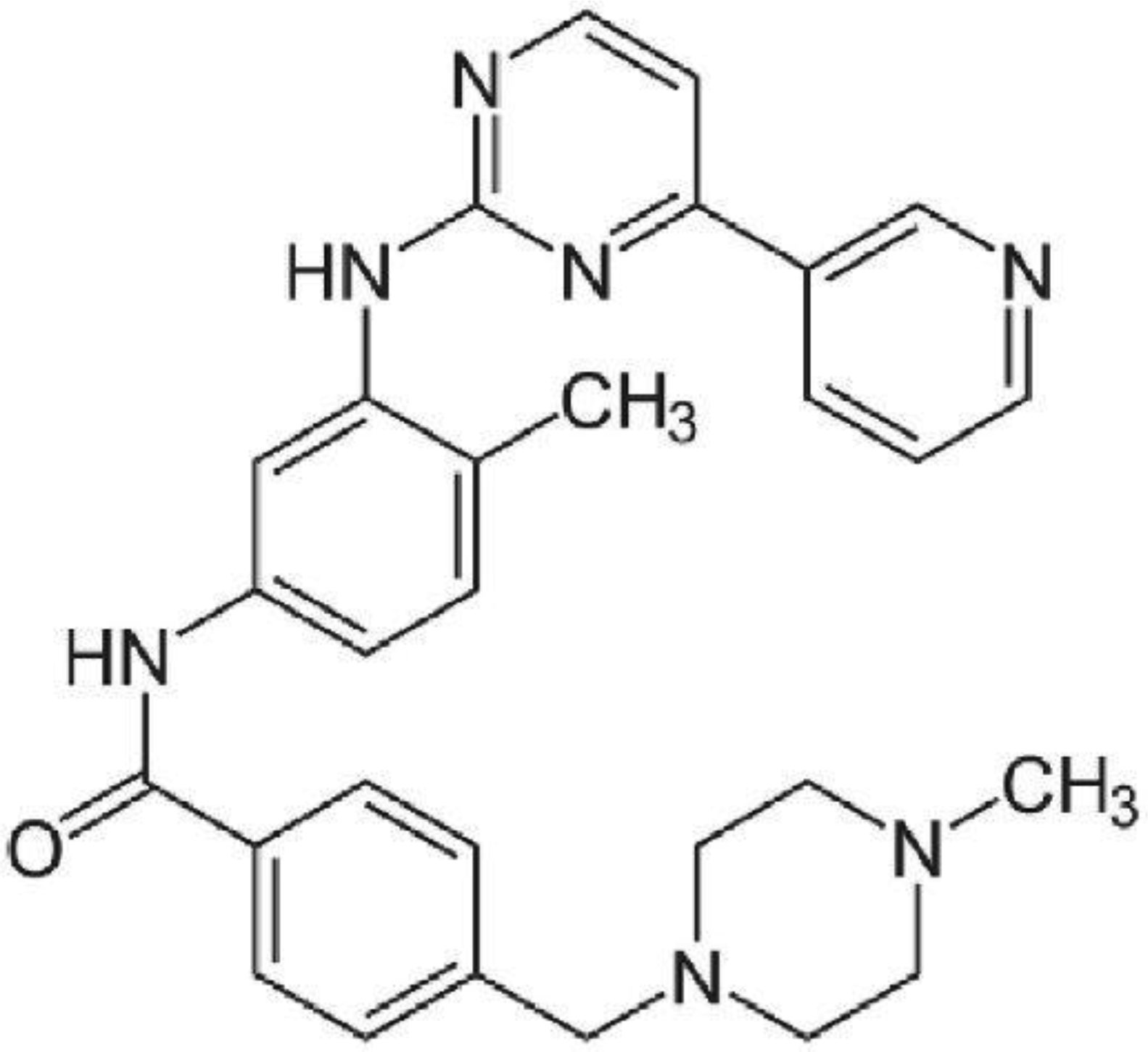
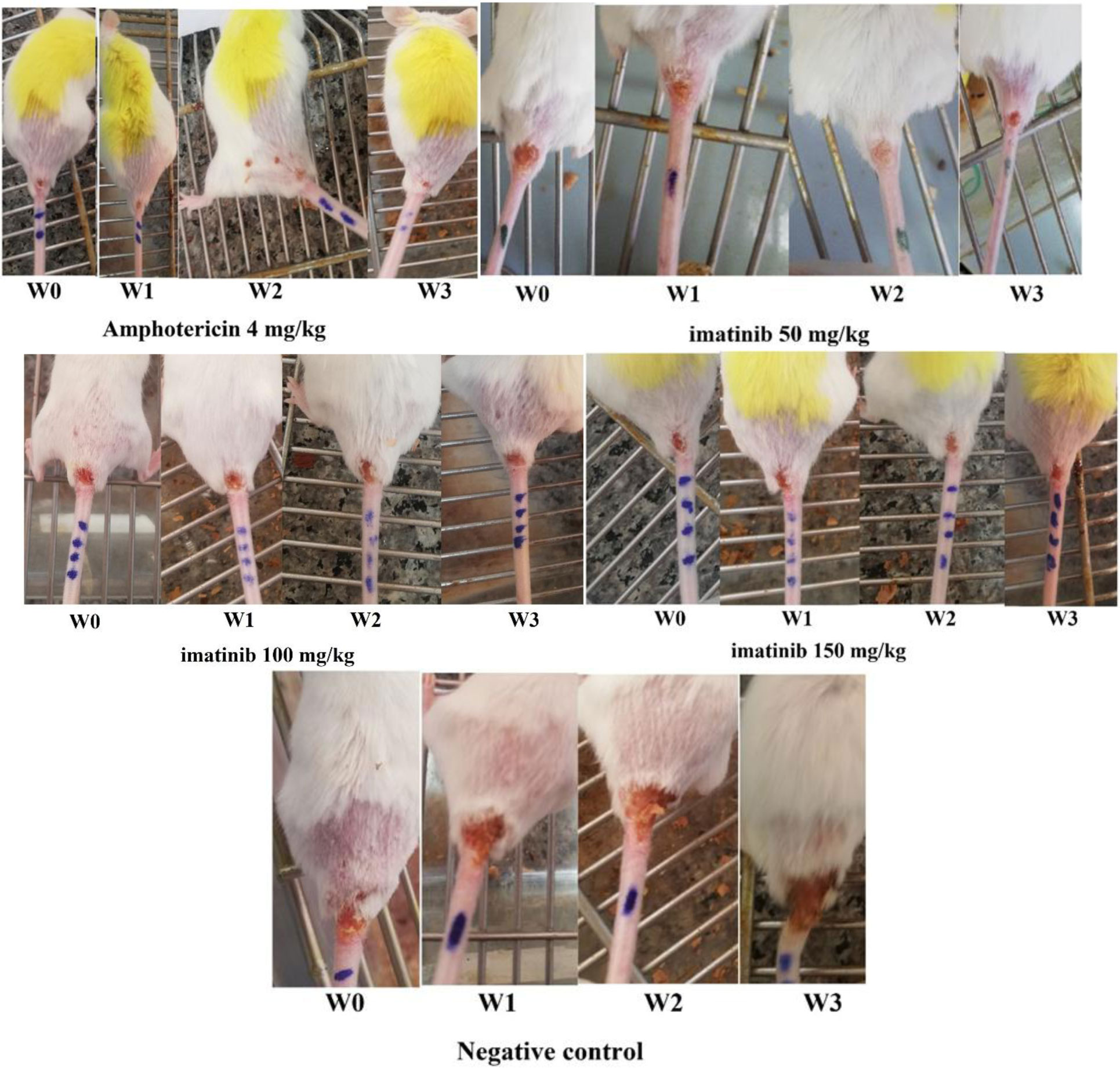
Leishmaniasis, as a tropical and subtropical disease, is endemic in more than 90 countries around the world. Today, cutaneous leishmaniasis (CL) that affects more than 1.5 million people per year lacks a definitive treatment approach. Imatinib is an anticancer drug that inhibits the abnormal function of Bcr-Abl due to its tyrosine kinase inhibitor, and that was the reason why the drug was tested for CL treatment because protein kinases are essential enzymes in the Leishmania genus. In this study, the L. major CL model of Balb/c mice was produced by injection of the cultured metacyclic form of parasite at the base of the tail. The possible recovery of CL ulcers and determination of the optimum dose of imatinib against Leishmania amastigotes were evaluated. A significant decrease was observed in mice treated with amphotericin B (positive control group) as well as imatinib 50 mg/kg compared to the unreceived drug, negative control group (P<0.05). This study could be promising in gaining insight into the potential of imatinib as an effective treatment approach against CL.
Citation: Mohsen Moslehi, Fatemeh Namdar, Mahsa Esmaeilifallah, Fariba Iraji, Bahareh Vakili, Fatemeh Sokhanvari, Seyed-Mohsen Hosseini, Faham Khamesipour, Zahra Sebghatollahi, Sayed-Hossein Hejazi. Study of therapeutic effect of different concentrations of imatinib on Balb/c model of cutaneous leishmaniasis[J]. AIMS Microbiology, 2020, 6(2): 152-161. doi: 10.3934/microbiol.2020010
Leishmaniasis, as a tropical and subtropical disease, is endemic in more than 90 countries around the world. Today, cutaneous leishmaniasis (CL) that affects more than 1.5 million people per year lacks a definitive treatment approach. Imatinib is an anticancer drug that inhibits the abnormal function of Bcr-Abl due to its tyrosine kinase inhibitor, and that was the reason why the drug was tested for CL treatment because protein kinases are essential enzymes in the Leishmania genus. In this study, the L. major CL model of Balb/c mice was produced by injection of the cultured metacyclic form of parasite at the base of the tail. The possible recovery of CL ulcers and determination of the optimum dose of imatinib against Leishmania amastigotes were evaluated. A significant decrease was observed in mice treated with amphotericin B (positive control group) as well as imatinib 50 mg/kg compared to the unreceived drug, negative control group (P<0.05). This study could be promising in gaining insight into the potential of imatinib as an effective treatment approach against CL.

| [1] |
Ghorbani M, Farhoudi R (2018) Leishmaniasis in humans: drug or vaccine therapy? Drug Des Devel Ther 12: 25-40. doi: 10.2147/DDDT.S146521

|
| [2] | Lindoso JA, Cunha MA, Queiroz IT, et al. (2016) Leishmaniasis–HIV coinfection: current challenges. HIV/AIDS 8: 147. |
| [3] |
Bahrami F, Harandi AM, Rafati S (2018) Biomarkers of cutaneous leishmaniasis. Front Cell Infect Microbiol 8: 222. doi: 10.3389/fcimb.2018.00222
|
| [4] |
Mendonça SC (2016) Differences in immune responses against Leishmania induced by infection and by immunization with killed parasite antigen: implications for vaccine discovery. Parasites Vectors 9: 492. doi: 10.1186/s13071-016-1777-x
|
| [5] |
Apostolopoulos N, Mitropoulou A, Thom N, et al. (2018) Update on therapy and prevention of canine leishmaniasis. Tierarztl Prax Ausg K Kleintiere Heimtiere 46: 315-322. doi: 10.15654/TPK-180089
|
| [6] | Kaiming B, Yuyang C, Songnian Z, et al. (2018) Current visceral leishmaniasis research: a research review to inspire future study. BioMed Res Intl 2018: 1-13. |
| [7] |
Norouzinezhad F, Ghaffari F, Norouzinejad A, et al. (2016) Cutaneous leishmaniasis in Iran: Results from an epidemiological study in urban and rural provinces. Asian Pac J Trop Biomed 6: 614-619. doi: 10.1016/j.apjtb.2016.05.005
|
| [8] |
Adriaensen W, Dorlo TPC, Guido V, et al. (2018) Immunomodulatory therapy of visceral leishmaniasis in human immunodeficiency virus-coinfected patients. Front Immunol 8: 1943. doi: 10.3389/fimmu.2017.01943
|
| [9] |
de Vries HJ, Reedijk SH, Schallig HD (2015) Cutaneous leishmaniasis: recent developments in diagnosis and management. Am J Clin Dermatol 16: 99-109. doi: 10.1007/s40257-015-0114-z
|
| [10] |
Alcântara LM, Ferreira TCS, Gadelha FR, et al. (2018) Challenges in drug discovery targeting TriTryp diseases with an emphasis on leishmaniasis. Int J Parasitol Drugs Drug Resist 8: 430-439. doi: 10.1016/j.ijpddr.2018.09.006
|
| [11] |
Hotez PJ, Bottazzi ME, Strych U (2016) New vaccines for the world's poorest people. Annu Rev Med 67: 405-417. doi: 10.1146/annurev-med-051214-024241
|
| [12] |
Moen MD, McKeage K, Plosker GL, et al. (2007) Imatinib: a review of its use in chronic myeloid leukaemia. Drugs 67: 299-320. doi: 10.2165/00003495-200767020-00010
|
| [13] |
O'Connell EM, Bennuru S, Steel C, et al. (2015) Targeting Filarial Abl-like Kinases: Orally Available, Food and Drug Administration–Approved Tyrosine Kinase Inhibitors Are Microfilaricidal and Macrofilaricidal. Int J Infect Dis 212: 684-693. doi: 10.1093/infdis/jiv065
|
| [14] |
Kesely KR, Pantaleo A, Turrini FM, et al. (2016) Inhibition of an erythrocyte tyrosine kinase with imatinib prevents Plasmodium falciparum egress and terminates parasitemia. PloS one 11. doi: 10.1371/journal.pone.0164895
|
| [15] |
Alvarez-Rueda N, Biron M, Le Pape (2009) Infectivity of Leishmania mexicana is associated with differential expression of protein kinase C-like triggered during a cell-cell contact. PLoS One 4. doi: 10.1371/journal.pone.0007581
|
| [16] |
Hodgson J (2001) ADMET—turning chemicals into drugs. Nat Biotechnol 19: 722-726. doi: 10.1038/90761
|
| [17] |
Potts RO, Guy RH (1992) Predicting skin permeability. Pharm Res 9: 663-669. doi: 10.1023/A:1015810312465
|
| [18] | Moslehi M, Namdar F, Esmaeilifallah M, et al. (2019) Evaluation of different concentrations of imatinib on the viability of Leishmania major: An In Vitro study. Adv Biomed Res 8. |
| [19] |
Ponte-Sucre A, Gamarro F, Dujardin JC, et al. (2017) Drug resistance and treatment failure in leishmaniasis: A 21st century challenge. PLoS Negl Trop Dis 11. doi: 10.1371/journal.pntd.0006052
|
| [20] |
Vakili B, Eslami M, Hatam GR, et al. (2018) Immunoinformatics-aided design of a potential multi-epitope peptide vaccine against Leishmania infantum. Int J Biol Macromol 120: 1127-1139. doi: 10.1016/j.ijbiomac.2018.08.125
|
| [21] |
Scott P, Novais FO (2016) Cutaneous leishmaniasis: immune responses in protection and pathogenesis. Nat Rev Immunol 16: 581-592. doi: 10.1038/nri.2016.72
|
| [22] |
Vakili B, Nezafat N, Zare B, et al. (2020) A new multi-epitope peptide vaccine induces immune responses and protection against Leishmania infantum in BALB/c mice. Med Microbiol Immunol 209: 69-79. doi: 10.1007/s00430-019-00640-7
|
| [23] |
Bekhit AA, El-Agroudy E, Helmy A, et al. (2018) Leishmania treatment and prevention: Natural and synthesized drugs. Eur J Med Chem 160: 229-244. doi: 10.1016/j.ejmech.2018.10.022
|
| [24] |
Vakili B, Nezafat N, Hatam GR, et al. (2018) Proteome-scale identification of Leishmania infantum for novel vaccine candidates: A hierarchical subtractive approach. Comput Biol Chem 72: 16-25. doi: 10.1016/j.compbiolchem.2017.12.008
|
| [25] |
Kennedy JA, Hobbs G (2018) Tyrosine kinase inhibitors in the treatment of chronic-phase CML: Strategies for frontline decision-making. Curr Hematol Malig Rep 13: 202-211. doi: 10.1007/s11899-018-0449-7
|
| [26] |
Katz N, Couto FFB, Araújo N (2013) Imatinib activity on Schistosoma mansoni. Mem Inst Oswaldo Cruz 108: 850-853. doi: 10.1590/0074-0276130207
|
| [27] |
Beckmann S, Long T, Scheld C, et al. (2014) Serum albumin and α-1 acid glycoprotein impede the killing of Schistosoma mansoni by the tyrosine kinase inhibitor Imatinib. Int J Parasitol Drugs Drug Resist 4: 287-295. doi: 10.1016/j.ijpddr.2014.07.005
|
| [28] |
Naula C, Parsons M, Mottram JC (2005) Protein kinases as drug targets in trypanosomes and Leishmania. Biochim Biophys Acta 1754: 151-159. doi: 10.1016/j.bbapap.2005.08.018
|
| [29] |
Handman E, Bullen DV (2002) Interaction of Leishmania with the host macrophage. Trends Parasitol 18: 332-334. doi: 10.1016/S1471-4922(02)02352-8
|
| [30] |
Sanderson L, Yardley V, Croft SL (2014) Activity of anticancer protein kinase inhibitors against Leishmania spp. J Antimicrob Chemother 69: 1888-1891. doi: 10.1093/jac/dku069
|
| [31] |
Matsushita M, Kawaguchi M (2018) Immunomodulatory effects of drugs for effective cancer immunotherapy. J Oncol 2018. doi: 10.1155/2018/8653489
|
| [32] |
Hahn T, Polanczyk MJ, Borodovsky A, et al. (2013) Use of anticancer drugs, mitocans, to enhance the immune responses against tumors. Curr Pharm Biotechnol 14: 357-376. doi: 10.2174/1389201011314030010
|
| [33] |
Al-Abdely HM, Graybill JR, Bocanegra R, et al. (1998) Efficacies of KY62 against Leishmania amazonensis and Leishmania donovani in experimental murine cutaneous leishmaniasis and visceral leishmaniasis. Antimicrob Agents Chemother 42: 2542-2548. doi: 10.1128/AAC.42.10.2542
|
| [34] | Dahiya M, Dureja H (2016) Central composite designed imatinib-loaded magnetic nanoparticles. Recent Pat Nanotechnol 6: 146-155. |
Figures(2) / Tables(2)

Mohsen Moslehi, Fatemeh Namdar, Mahsa Esmaeilifallah, Fariba Iraji, Bahareh Vakili, Fatemeh Sokhanvari, Seyed-Mohsen Hosseini, Faham Khamesipour, Zahra Sebghatollahi, Sayed-Hossein Hejazi. Study of therapeutic effect of different concentrations of imatinib on Balb/c model of cutaneous leishmaniasis[J]. AIMS Microbiology, 2020, 6(2): 152-161. doi: 10.3934/microbiol.2020010







 DownLoad:
DownLoad: