Citation: Priscilla Charmaine Kwade, Benjamin Kweku Lugu, Sadia Lukman, Carl Edem Quist, Jianxun Chu. Farmers’ attitude towards the use of genetically modified crop technology in Southern Ghana: The mediating role of risk perception[J]. AIMS Agriculture and Food, 2019, 4(4): 833-853. doi: 10.3934/agrfood.2019.4.833

| [1] | Kwade C, Gadabu A, Addo V, et al. (2018) Genetic engineering development and acceptance in Sub Saharan Africa: Stakeholders and public opinion about GMO. J Appl Agric Econ Policy Anal 1: 22-30. |
| [2] | Jayne T, Govereh J, Mwanaumo A, et al. (2002) False promise or false premise? The experience of food and input market reform in Eastern and Southern Africa. World Dev 30: 1967-1985 |
| [3] |
Sarcheshmeh EE, Bijani M, Sadighi H (2018) Technology in Society Adoption behavior towards the use of nuclear technology in agriculture: A causal analysis. Technol Soc 55: 175-182. Available from: https://doi.org/10.1016/j.techsoc.2018.08.001. doi: 10.1016/j.techsoc.2018.08.001

|
| [4] | Bailey R, Willoughby R, Grzywacz D (2014) On trial: Agricultural biotechnology in Africa. Energy Environ Resour 1-27. |
| [5] |
Qaim M, Kouser S (2013) Genetically modified crops and food security. PloS one 8: e64879. doi: 10.1371/journal.pone.0064879
|
| [6] | Séralini GE, Clair E, Mesnage R, et al. (2012) RETRACTED: Long term toxicity of a Roundup herbicide and a Roundup-tolerant genetically modified maize. Food Chem Toxicol 4221-4231. Available from: http://dx.doi.org/10.1016/j.fct.2012.08.005. |
| [7] |
Adenle AA, Morris EJ, Parayil G (2013) Status of development, regulation and adoption of GM agriculture in Africa: Views and positions of stakeholder groups. Food Policy 43: 159-166. Available from: https://doi.org/10.1016/j.foodpol.2013.09.006. doi: 10.1016/j.foodpol.2013.09.006
|
| [8] | Rosegrant MW, Koo J, Cenacchi N, et al. (2014) Food security in a world of natural resource scarcity: The role of agricultural technologies. In: International Food Policy Research Institute, Washington, DC, USA. |
| [9] | Rogers EM, Singhal A, Quinlan MM (2003) Diffusion of innovations/Everett M. Rogers. NY: Simon and Schuster, 576. |
| [10] |
Costa-Font M, Gil JM, Traill WB (2008) Consumer acceptance, valuation of and attitudes towards genetically modified food: Revies and implications for food policy. Food Policy 33: 99-111. doi: 10.1016/j.foodpol.2007.07.002
|
| [11] |
Mannan S, Nordin SM, Rafik-Galea S, et al. (2017) The ironies of new innovation and the sunset industry: Diffusion and adoption. J Rural Studies 55: 316-322. doi: 10.1016/j.jrurstud.2017.07.015
|
| [12] |
Ueland Ø, Gunnlaugsdottir H, Holm F, et al. (2012) State of the art in benefit-risk analysis: Consumer perception. Food Chem Toxicol 50: 67-76. doi: 10.1016/j.fct.2011.06.006
|
| [13] | Rogers E (2003) Diffusion of innovations. New York: A Division of Simon and Schuster. |
| [14] | Ghane F, Samah BA, Ahmad A, et al. (2011) The role of social influence and innovation characteristics in the adoption of Integrated Pest Management (IPM) practices by paddy farmers in Iran. In: International Conference on Social Science and Humanity-IPEDR Singapore. |
| [15] | Peshin R, Vasanthakumar J, Kalra R (2009) Diffusion of innovation theory and integrated pest management. Integr Pest Manage: Dissemination Impact: 1-29. Springer, Dordrecht. Available from: https://doi.org/10.1007/978-1-4020-8990-9. |
| [16] |
Robertson MJ, Llewellyn RS, Mandel R, et al. (2012) Adoption of variable rate fertiliser application in the Australian grains industry: Status, issues and prospects. Precis Agric 13: 181-199. doi: 10.1007/s11119-011-9236-3
|
| [17] |
Hirunyawipada T, Paswan AK (2006) Consumer innovativeness and perceived risk: Implications for high technology product adoption. J Consum Mark 23: 182-198. doi: 10.1108/07363760610674310
|
| [18] | Taylor JW (1974) The role of risk in consumer behavior: A comprehensive and operational theory of risk taking in consumer behavior. J Mark 38: 54-60. |
| [19] |
Gupta N, Fischer AR, Frewer LJ (2012) Socio-psychological determinants of public acceptance of technologies: A review. Public Understanding Sci 21: 782-795. Available from: http://dx.doi.org/10.1177/0963662510392485. doi: 10.1177/0963662510392485
|
| [20] | Kim RB (2012) Consumer attitude of risk and benefits toward genetically modified (GM) foods in South Korea: Implications for food policy. Eng Econ 23: 189-199. Available from: http://dx.doi.org/10.5755/j01.ee.23.2.1548. |
| [21] |
Gonçalves ME (2006) Risk and the governance of innovation in Europe: An introduction. Technol Forecasting Social Change 73: 1-12. doi: 10.1016/j.techfore.2005.06.008
|
| [22] | Sjöberg L (2008) Genetically modified food in the eyes of the public and experts. Risk Manage 10: 168-193. Available from: http://dx.doi.org/10.1057/rm.2008.2. |
| [23] | Miller DM, Meek F (2004) Cost and efficacy comparison of integrated pest management strategies with monthly spray insecticide applications for German cockroach (Dictyoptera: Blattellidae) control in public housing. J Econ Entomol 97: 559-569. |
| [24] | Sjakir M, Awang AH, Azima AM, et al. (2015) Learning and technology adoption impacts on farmer's productivity. Mediterr J Soc Sci 6: 126. |
| [25] | Kolodinsky JM, Hogarth JM, Hilgert MA (2004) The adoption of electronic banking technologies by US consumers. Int J Bank Market 22: 238-259. |
| [26] |
Ronteltap A, Van Trijp JCM, Renes RJ, et al. (2007) Consumer acceptance of technology-based food innovations: Lessons for the future of nutrigenomics. Appetite 49: 1-17. doi: 10.1016/j.appet.2007.02.002
|
| [27] | Gaskell G, Bauer MW, Durant J, et al. (1999) Worlds apart? The reception of genetically modified foods in Europe and the US. Science 285: 384-387. |
| [28] | Talebian A, Mishra S (2018) Predicting the adoption of connected autonomous vehicles: A new approach based on the theory of diffusion of innovations. Transp Res Part C: Emerging Technol 95: 363-380. |
| [29] | Lagnaoui A, Santi E, Santucci F (2004) Strategic communication for integrated pest management. Paper first presented at the Annual Conference of the International Association for Impact Assessment in Vancouver, Canada (IAIA 2004) under the conference theme of Public Involvement and Risk Management. Available from: http://siteresources.worldbank.org/ extdevcomsusdevt/Resources/ipmcommfinal.pdf. |
| [30] |
Bardin B, Perrissol S, Facca L, et al. (2017) From risk perception to information selection… And not the other way round: Selective exposure mechanisms in the field of genetically modified organisms. Food Quality Preference 58: 10-17. Available from: https://doi.org/10.1016/j.foodqual.2016.12.015. doi: 10.1016/j.foodqual.2016.12.015
|
| [31] |
Swartz DG, Strand IE (1981) Avoidance costs associated with imperfect information: The case of kepone. Land Econ 57: 139-150. doi: 10.2307/3145782
|
| [32] |
De Barcellos MD, Kügler JO, Grunert KG, et al. (2010) European consumers' acceptance of beef processing technologies: A focus group study. Innovative Food Sci Emerging Technol 11: 721-732. Available from: https://doi.org/10.1016/j.ifset.2010.05.003. doi: 10.1016/j.ifset.2010.05.003
|
| [33] | Verbeke W, Ward RW (2001) A fresh meat almost ideal demand system incorporating negative TV press and advertising impact. Agric Econ 25: 359-374. |
| [34] | Rock J (2013) Food sovereignty and the Anti-GMO movement in Ghana. Modern Ghana. Available from: http://www.modernghana.com/news/487275/1/food-sovereignty-and-the-antigmo-movement-in-ghan.html. |
| [35] |
Mabaya E, Fulton J, Simiyu-Wafukho S, et al. (2015) Factors influencing adoption of genetically modified crops in Africa. Dev South Afr 32: 577-591. Available from: https://doi.org/10.1080/0376835X.2015.1044078. doi: 10.1080/0376835X.2015.1044078
|
| [36] | Ammar RAR, Shahrina N, Shamsuri S, et al. (2013) Communication management between organizations in a subsidized market: The Case of Paddy Fertilizer in Malaysia. Asian J Res Soc Sci Humanit 3: 1-16. |
| [37] |
Dibden J, Gibbs D, Cocklin C (2013) Framing GM crops as a food security solution. J Rural Stud 29: 59-70. Available from: https://doi.org/10.1016/j.jrurstud.2011.11.001. doi: 10.1016/j.jrurstud.2011.11.001
|
| [38] |
Scott S (2008) East Asian experience in environmental governance: Response in a rapidly developing region‐edited by Zafar Adeel. Geogr Res 46: 126-127. doi: 10.1111/j.1745-5871.2007.00498.x
|
| [39] |
Horst M, Kuttschreuter M, Gutteling JM (2007) Perceived usefulness, personal experiences, risk perception and trust as determinants of adoption of e-government services in the Netherlands. Comput Human Behav 23: 1838-1852. Available from: https://doi.org/10.1016/j.chb.2005.11.003. doi: 10.1016/j.chb.2005.11.003
|
| [40] |
Featherman MS, Pavlou PA (2003) Predicting e-services adoption: A perceived risk facets perspective. Int J Human Comput Studies 59: 451-474. doi: 10.1016/S1071-5819(03)00111-3
|
| [41] | Stoner KA, Sawyer AJ, Shelton AM (1986) Constraints to the implementation of IPM programs in the USA: A course outline. Agric Ecosyst Environ 17: 253-268. |
| [42] |
Vilella-Vila M, Costa-Font J (2008) Press media reporting effects on risk perceptions and attitudes towards genetically modified (GM) food. J Socio-Econo 37: 2095-2106. Available from: https://doi.org/10.1016/j.socec.2008.04.006. doi: 10.1016/j.socec.2008.04.006
|
| [43] |
Bearth A, Siegrist M (2016) Are risk or benefit perceptions more important for public acceptance of innovative food technologies: A meta-analysis. Trends Food Sci Technol 49: 14-23. Available from: https://doi.org/10.1016/j.tifs.2016.01.003. doi: 10.1016/j.tifs.2016.01.003
|
| [44] |
Vänninen I, Siipi H, Keskitalo M, et al. (2009) Ethical compatibility of GM crops with intrinsic and extrinsic values of farms: A review. Open Ethics J 3: 104-117. doi: 10.2174/1874761200903030104
|
| [45] | Fishbein M, Ajzen I (1975a) Belief, attitude, intention, and behavior: An introduction to theory and research. Reading, MA: Addison-Wesley. |
| [46] | Ajzen I (1991) The theory of planned behavior. Organ Behav Human Decis Processes 50: 179-211. |
| [47] | Burke J (2014) Biotechnology crop adoption: Potential and challenges of genetically improved crops. Encycl Agric Food Syst: 69-93. |
| [48] |
Nordin SM, Noor SM, bin Md Saad MS (2014) Innovation diffusion of new technologies in the malaysian paddy fertilizer industry. Procedia-Soc Behav Sci 109: 768-778. doi: 10.1016/j.sbspro.2013.12.542
|
| [49] |
Valente M, Chaves C (2018) Perceptions and valuation of GM food: A study on the impact and importance of information provision. J Cleaner Prod 172: 4110-4118. Available from: https://doi.org/10.1016/j.jclepro.2017.02.042. doi: 10.1016/j.jclepro.2017.02.042
|
| [50] |
Aerni P (2005) Stakeholder attitudes towards the risks and benefits of genetically modified crops in South Africa. Environ Sci Policy 8: 464-476. Available from: https://doi.org/10.1016/j.envsci.2005.07.001. doi: 10.1016/j.envsci.2005.07.001
|
| [51] | Fishbein M, Ajzen I (1975b) Belief, attitude, intention, and behavior: An introduction to theory and research. Reading, MA: Addison-Wesley. |
| [52] |
Pardo R, Midden C, Miller JD (2002) Attitudes toward biotechnology in the European Union. J Biotechnol 98: 9-24. doi: 10.1016/S0168-1656(02)00082-2
|
| [53] | Hair JF, Black WC, Babin BJ (2010) Multivariate data analysis (7 Eds.). Upper Saddle River, NJ: Pearson Prentice Hall. |
| [54] |
Fornell C, Larcker D (1981) Evaluating structural equation models with unobservable variables and measurement errors. J Mark Res 18: 39-50. doi: 10.1177/002224378101800104
|
| [55] |
Chiu CM, Wang ET (2008) Understanding Web-based learning continuance intention: The role of subjective task value. Inf Manage 45: 194-201. doi: 10.1016/j.im.2008.02.003
|
| [56] | Bagozzi RP, Yi Y, Phillips LW (1991) Assessing construct validity in organizational research. Administrative Sci Q: 421-458. |
| [57] |
Arts JWC, Frambach RT, Bijmolt THA (2011) Generalizations on consumer innovation adoption: A meta-analysis on drivers of intention and behavior. Int J Res Mark 28: 134-144. Available from: https://doi.org/10.1016/j.ijresmar.2010.11.002. doi: 10.1016/j.ijresmar.2010.11.002
|
| [58] |
Zhang Y, Jing L, Bai Q, et al. (2018) Application of an integrated framework to examine Chinese consumers' purchase intention toward genetically modified food. Food Quality Preference 65: 118-128. Available from: https://doi.org/10.1016/j.foodqual.2017.11.001. doi: 10.1016/j.foodqual.2017.11.001
|
| [59] |
Zhao X, Lynch Jr JG, Chen Q (2010) Reconsidering Baron and Kenny: Myths and truths about mediation analysis. J Consum Res 37: 197-206. Available from: https://doi.org/10.1086/651257. doi: 10.1086/651257
|
| [60] | Gyau A, Voss J, Spiller A, et al. (2009) Farmer acceptance of genetically modified seeds in Germany: Results of a cluster analysis. Int Food Agribusiness Manage Rev 12(1030-2016-82757): 1-20. |
| [61] |
Costa-Font M, Gil JM (2009) Structural equation modelling of consumer acceptance of genetically modified (GM) food in the Mediterranean Europe: A cross country study. Food Qual Preference 20: 399-409. Available from: https://doi.org/10.1016/j.foodqual.2009.02.011. doi: 10.1016/j.foodqual.2009.02.011
|
| [62] |
Chen MF, Li HL (2007) The consumer's attitude toward genetically modified foods in Taiwan. Food Qual Preference 18: 662-674. doi: 10.1016/j.foodqual.2006.10.002
|
| [63] | Bredahl L, Grunert KG, Frewer LJ (1998) Consumer attitudes and decisionmaking with regard to genetically engineered food products. A review of the literature and a presentation of models for future research. J Consum Policy 21: 251-277. |
| [64] |
Weick CW, Walchli SB (2002) Genetically engineered crops and foods: Back to the basics of technology diffusion. Technol Soc 24: 265-283. doi: 10.1016/S0160-791X(02)00008-8
|
| [65] |
Nelson CH (2001) Risk perception, behavior and consumer response to genetically modified organisms, toward and understanding American and European pubic reaction. Am Behav Sci 44: 1371-1388. doi: 10.1177/00027640121956737
|
| [66] |
Martinez-Poveda A, Brugarolas Molla-Bauza M, del Campo Gomis FJ, et al. (2009) Consumer-perceived risk model for the introduction of genetically modified food in Spain. Food Policy 34: 519-528. doi: 10.1016/j.foodpol.2009.08.001
|
| [67] | Amin L, Mahadi Z, Samian AL, et al. (2013a) Risk perception towards food safety issues: GM foods versus non-GM foods. J Food Agric Environ 11: 28-35. |
| [68] | Onyango B, Govindasamy R (2004) Measuring US consumer preferences for genentically modified foods using choice modelling experiments: The role of price, product benefits and technology. In: American Agricultural Economics Association annual meeting, Colorado. |
| [69] | Amin L, Mahadi Z, Samian AL, et al. (2013b) Risk perception towards food safety issues: GM foods versus non-GM foods. J Food Agric Environ 11: 28-35. |
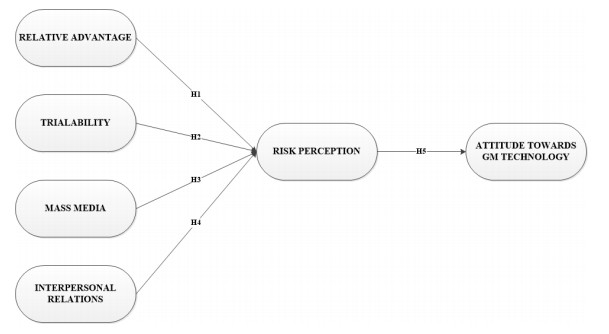
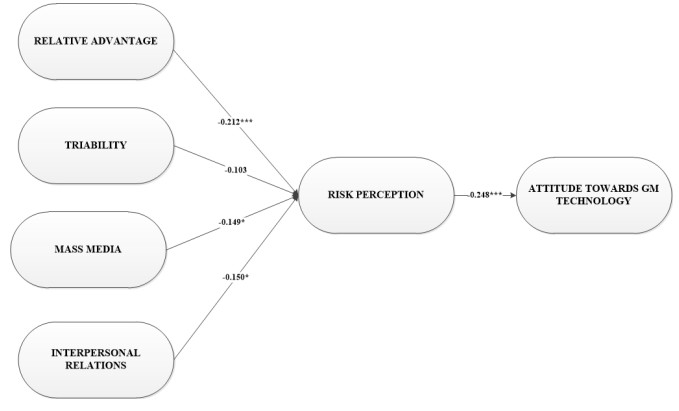
Figures(2) / Tables(6)

Priscilla Charmaine Kwade, Benjamin Kweku Lugu, Sadia Lukman, Carl Edem Quist, Jianxun Chu. Farmers’ attitude towards the use of genetically modified crop technology in Southern Ghana: The mediating role of risk perception[J]. AIMS Agriculture and Food, 2019, 4(4): 833-853. doi: 10.3934/agrfood.2019.4.833







 DownLoad:
DownLoad: