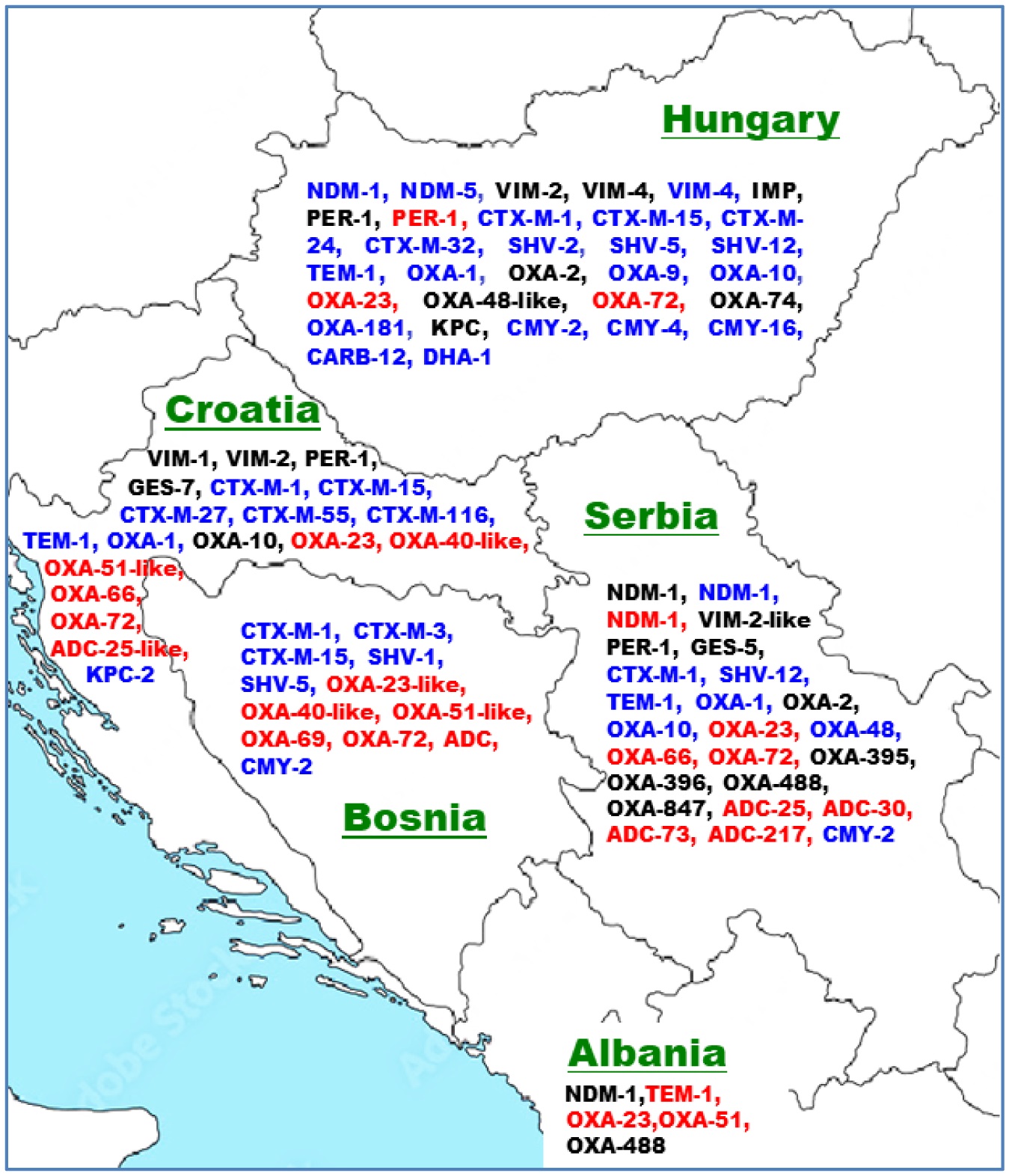
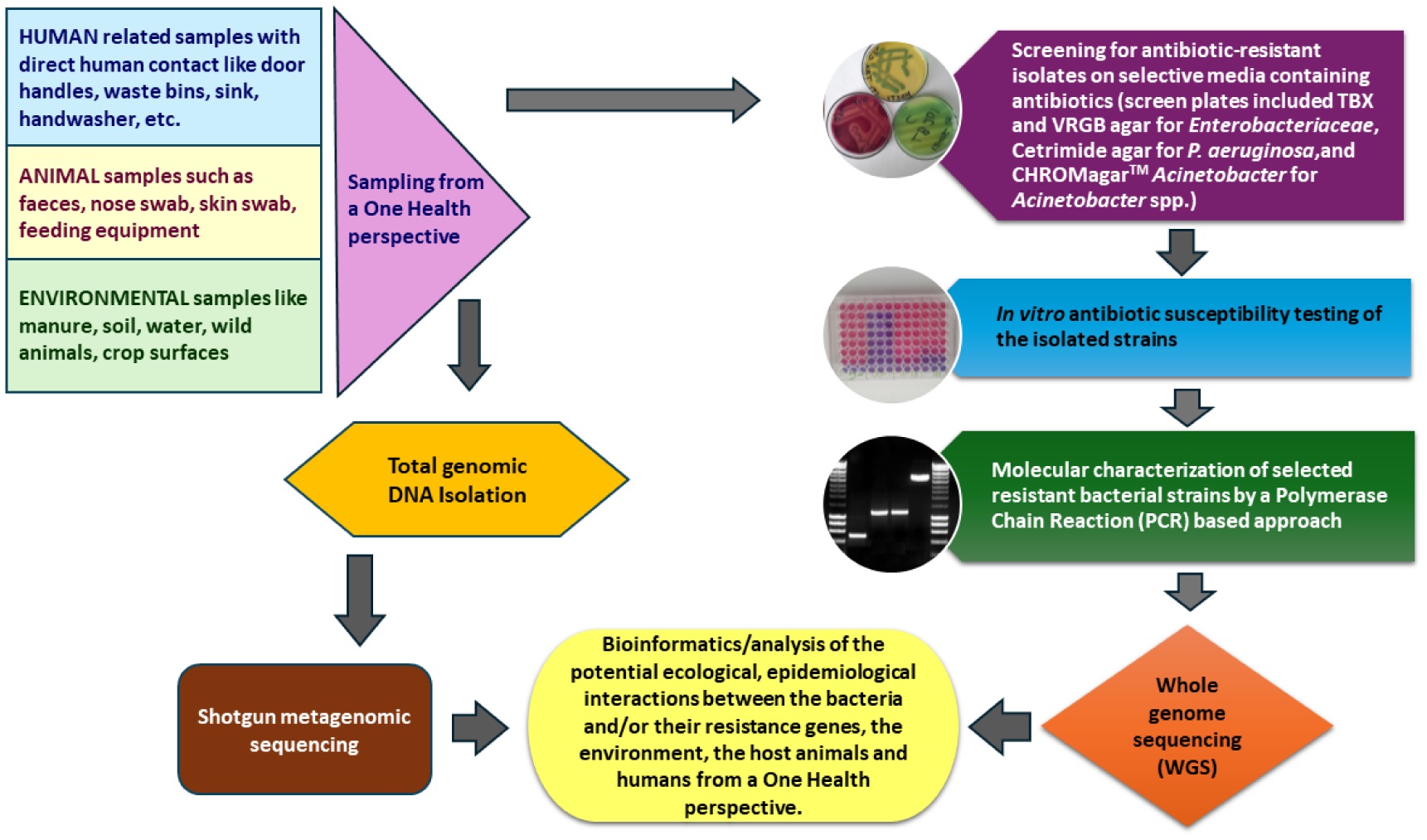
An increasing rate of antibiotic resistance (AR) has been observed in the Gram-negative bacteria A. baumannii, P. aeruginosa, and E. coli in the human, environmental, and food animal domains worldwide, thus posing a serious global health challenge. Acquired AR genes of these species were overviewed from selected Western Balkans countries together with those from the European Union member states Croatia and Hungary. The AR determinants published from Albania, Bosnia-Herzegovina, Serbia, and Croatia included diverse acquired β-lactamase genes, with several of them possessing carbapenemase activity, such as blaVIM, blaNDM, blaKPC, blaOXA-23, blaOXA-66, and blaOXA-72. Furthermore, acquired aminoglycoside, chloramphenicol, fosfomycin, tetracycline, sulfonamide, quinolone, and/or colistin resistance determinants were detected in the three domains of the One Health approach. The in vitro AR profile of representative isolates have also been overviewed. Multidrug-resistant P. aeruginosa isolates of the ST235 high-risk clone were mainly reported within clinical settings. The distribution of the E. coli ST131 and A. baumannii ST2 high-risk clones in both clinical and environmental settings highlight their adaptability and effective dissemination. Systematic infection control practices are advised to combat the spread of antibiotic resistance, and further research from a One Health perspective is encouraged into its emergence and dissemination.
Citation: Chioma Lilian Ozoaduche, Katalin Posta, Balázs Libisch, Ferenc Olasz. Acquired antibiotic resistance of Pseudomonas spp., Escherichia coli and Acinetobacter spp. in the Western Balkans and Hungary with a One Health outlook[J]. AIMS Microbiology, 2025, 11(2): 436-461. doi: 10.3934/microbiol.2025020
An increasing rate of antibiotic resistance (AR) has been observed in the Gram-negative bacteria A. baumannii, P. aeruginosa, and E. coli in the human, environmental, and food animal domains worldwide, thus posing a serious global health challenge. Acquired AR genes of these species were overviewed from selected Western Balkans countries together with those from the European Union member states Croatia and Hungary. The AR determinants published from Albania, Bosnia-Herzegovina, Serbia, and Croatia included diverse acquired β-lactamase genes, with several of them possessing carbapenemase activity, such as blaVIM, blaNDM, blaKPC, blaOXA-23, blaOXA-66, and blaOXA-72. Furthermore, acquired aminoglycoside, chloramphenicol, fosfomycin, tetracycline, sulfonamide, quinolone, and/or colistin resistance determinants were detected in the three domains of the One Health approach. The in vitro AR profile of representative isolates have also been overviewed. Multidrug-resistant P. aeruginosa isolates of the ST235 high-risk clone were mainly reported within clinical settings. The distribution of the E. coli ST131 and A. baumannii ST2 high-risk clones in both clinical and environmental settings highlight their adaptability and effective dissemination. Systematic infection control practices are advised to combat the spread of antibiotic resistance, and further research from a One Health perspective is encouraged into its emergence and dissemination.

| [1] |
Breijyeh Z, Jubeh B, Karaman R (2020) Resistance of Gram-Negative bacteria to current antibacterial agents and approaches to resolve it. Molecules 25: 1340. https://doi.org/10.3390/molecules25061340 
|
| [2] | WHO Regional Office for Europe/European Centre for Disease Prevention and ControlAntimicrobial resistance surveillance in Europe 2022–2020 data (2022). Available from: https://www.ecdc.europa.eu/en/publications-data/antimicrobial-resistance-surveillance-europe-2022-2020-data |
| [3] | Central Asian and European surveillance of antimicrobial resistance: Annual report 2019. Available from: https://www.who.int/europe/groups/central-asian-and-european-surveillance-of-antimicrobial-resistance-(caesar) |
| [4] |
Bharadwaj A, Rastogi A, Pandey S, et al. (2022) Multidrug-resistant bacteria: Their mechanism of action and prophylaxis. Biomed Res Int 2022: 5419874. https://doi.org/10.1155/2022/5419874
|
| [5] |
Abduzaimovic A, Aljicevic M, Rebic V, et al. (2016) Antibiotic resistance in urinary isolates of Escherichia coli. Mater Sociomed 28: 416-419. https://doi.org/10.5455/msm.2016.28.416-419
|
| [6] |
Kot B (2019) Antibiotic resistance among uropathogenic Escherichia coli. Pol J Microbiol 68: 403-415. https://doi.org/10.33073/pjm-2019-048
|
| [7] |
Libisch B (2013) Molecular typing methods for the genus Pseudomonas. Molecular Typing in Bacterial Infections . Totowa: Humana Press 407-429. https://doi.org/10.1007/978-1-62703-185-1_24
|
| [8] |
Radovanovic RS, Savic NR, Ranin L, et al. (2020) Biofilm production and antimicrobial resistance of clinical and food isolates of Pseudomonas spp. Curr Microbiol 77: 4045-4052. https://doi.org/10.1007/s00284-020-02236-4
|
| [9] |
Schwartz T, Volkmann H, Kirchen S, et al. (2006) Real-time PCR detection of Pseudomonas aeruginosa in clinical and municipal wastewater and genotyping of the ciprofloxacin-resistant isolates. FEMS Microbiol Ecol 57: 158-167. https://doi.org/10.1111/j.1574-6941.2006.00100.x
|
| [10] | European Centre for Disease Prevention and Control and World Health OrganizationAntimicrobial resistance surveillance in Europe 2023–2021 data (2023). Available from: https://www.ecdc.europa.eu/en/publications-data/antimicrobial-resistance-surveillance-europe-2023-2021-data |
| [11] |
Manyi-Loh C, Mamphweli S, Meyer E, et al. (2018) Antibiotic use in agriculture and its consequential resistance in environmental sources: potential public health implications. Molecules 23: 795. https://doi.org/10.3390/molecules23040795
|
| [12] |
Garcia-Alvarez L, Dawson S, Cookson B, et al. (2012) Working across the veterinary and human health sectors. J Antimicrob Chemother 67: i37-i49. https://doi.org/10.1093/jac/dks206
|
| [13] | Regulation (EU) 2019/6 of the European parliament and of the council of 11 December 2018 on veterinary medicinal products and repealing Directive 2001/82/EC (Text with EEA relevance). OJEU 43-167. Available from: http://data.europa.eu/eli/reg/2019/6/oj |
| [14] | U.S. Food and Drug AdministrationAntimicrobial Resistance (2025). Available from: https://www.fda.gov/animal-veterinary/safety-health/antimicrobial-resistance |
| [15] |
Roth N, Käsbohrer A, Mayrhofer S, et al. (2019) The application of antibiotics in broiler production and the resulting antibiotic resistance in Escherichia coli: A global overview. Poult Sci 98: 1791-804. https://doi.org/10.3382/ps/pey539
|
| [16] |
Xu C, Kong L, Gao H, et al. (2022) A review of current bacterial resistance to antibiotics in food animals. Front Microbiol 13: 822689. https://doi.org/10.3389/fmicb.2022.822689
|
| [17] |
Larsson DGJ, Flach CF (2022) Antibiotic resistance in the environment. Nat Rev Microbiol 20: 257-269. https://doi.org/10.1038/s41579-021-00649-x
|
| [18] |
de Lorenzo Gilsanz FJ, Flores Martos R, Mora Rosado S (2025) Unpacking vulnerability among migrant minors in the EU: A case study of vulnerability among migrant minors in Spain. Int J Soc Welf 34: e12717. https://doi.org/10.1111/ijsw.12717
|
| [19] | Berishaj V, Schwadere G, Arning A, et al. (2024) Biodiversity. The case of the Western Balkans, Europe: EuroNatur Foundation. Available from: https://www.euronatur.org/fileadmin/docs/umweltpolitik/Biodiversitaet/EuroNatur_Biodiversity_Report_211124.pdf. |
| [20] |
Foti M, Mascetti A, Fisichella V, et al. (2017) Antibiotic resistance assessment in bacteria isolated in migratory Passeriformes transiting through the Metaponto territory (Basilicata, Italy). Avian Res 8: 26. https://doi.org/10.1186/s40657-017-0085-2
|
| [21] |
Elsohaby I, Samy A, Elmoslemany A, et al. (2021) Migratory wild birds as a potential disseminator of antimicrobial-resistant bacteria around Al-Asfar Lake, Eastern Saudi Arabia. Antibiotics 10: 260. https://doi.org/10.3390/antibiotics10030260
|
| [22] | Rahman A, Chowdhury Md.SR, Hossain H, et al. (2024) Identification of virulence genes and multidrug resistance in Shiga-toxin producing Escherichia coli (STEC) from migratory and captive wild birds. Pak Vet J 44: 1120-1130. http://dx.doi.org/10.29261/pakvetj/2024.264 |
| [23] |
Magiorakos AP, Srinivasan A, Carey RB, et al. (2012) Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect 18: 268-281. https://doi.org/10.1111/j.1469-0691.2011.03570.x
|
| [24] | Centers for Disease Control and Prevention (U.S.), National Center for Emerging Zoonotic and Infectious Diseases (U.S.), Division of Healthcare Quality Promotion, and Antibiotic Resistance Coordination and Strategy UnitAntibiotic resistance threats in the United States, 2019 (2019). http://dx.doi.org/10.15620/cdc:82532 |
| [25] |
Tsanova D (2020) The Balkan countries in global burden of diseases studies. Arch Balk Medical Union 55: 684-690. https://doi.org/10.31688/ABMU.2020.55.4.16
|
| [26] |
Peykov S, Strateva T (2023) Whole-Genome Sequencing-Based resistome analysis of nosocomial multidrug-resistant non-fermenting Gram-Negative pathogens from the Balkans. Microorganisms 11: 651. https://doi.org/10.3390/microorganisms11030651
|
| [27] | Jonas OB, Irwin A, Berthe FCJ, et al. Drug-resistant infections: A threat to our economic future (Vol. 2): FInal report (English) (2017). Available from: http://documents.worldbank.org/curated/en/323311493396993758/final-report. |
| [28] |
Lepsanovic Z, Libisch B, Tomanovic B, et al. (2008) Characterisation of the first VIM metallo-β-lactamase-producing Pseudomonas aeruginosa clinical isolate in Serbia. Acta Microbiol Immunol Hung 55: 447-454. https://doi.org/10.1556/AMicr.55.2008.4.9
|
| [29] | Libisch B, Poirel L, Lepsanovic Z, et al. (2008) Identification of PER-1 extended-spectrum β-lactamase producing Pseudomonas aeruginosa clinical isolates of the international clonal complex CC11 from Hungary and Serbia. FEMSIM 54: 330-338. https://doi.org/10.1111/j.1574-695X.2008.00483.x |
| [30] |
Jovcic B, Lepsanovic Z, Suljagic V, et al. (2011) Emergence of NDM-1 metallo-β-lactamase in Pseudomonas aeruginosa clinical isolates from Serbia. Antimicrob Agents Chemother 55: 3929-3931. https://doi.org/10.1128/AAC.00226-11
|
| [31] |
Jovčić B, Lepšanović Z, Begović J, et al. (2014) Two copies of blaNDM-1 gene are present in NDM-1 producing Pseudomonas aeruginosa isolates from Serbia. Antonie Van Leeuwenhoek 105: 613-618. https://doi.org/10.1007/s10482-013-0094-z
|
| [32] |
Kabic J, Fortunato G, Vaz-Moreira I, et al. (2023) Dissemination of metallo-β-lactamase-producing Pseudomonas aeruginosa in Serbian hospital settings: Expansion of ST235 and ST654 clones. Int J Mol Sci 24: 1519. https://doi.org/10.3390/ijms24021519
|
| [33] |
Kabic J, Novovic K, Kekic D, et al. (2022) Comparative genomics and molecular epidemiology of colistin-resistant Acinetobacter baumannii. Comput Struct Biotechnol J 21: 574-585. https://doi.org/10.1016/j.csbj.2022.12.045
|
| [34] |
Tafaj S, Gona F, Rodrigues CF, et al. (2020) Whole-Genome Sequences of two NDM-1-Producing Pseudomonas aeruginosa strains isolated in a clinical setting in Albania in 2018. Microbiol Resour Announc 9. https://doi.org/10.1128/MRA.01291-19
|
| [35] |
Dortet L, Bonnin RA, Bernabeu S, et al. (2016) First Occurrence of OXA-72-Producing Acinetobacter baumannii in Serbia. Antimicrob Agents Chemother 60: 5724-5730. https://doi.org/10.1128/AAC.01016-16
|
| [36] |
Gajic I, Jovicevic M, Milic M, et al. (2021) Clinical and molecular characteristics of OXA-72-producing Acinetobacter baumannii ST636 outbreak at a neonatal intensive care unit in Serbia. J Hosp Infect 112: 54-60. https://doi.org/10.1016/j.jhin.2021.02.023
|
| [37] | Abdelbary MMH, Prod'hom G, Greub G, et al. (2017) Draft genome sequences of two Carbapenemase-Producing Acinetobacter baumannii clinical strains isolated from Albanian and Togolese Patients. Genome Announc 5: e00115-e00117. https://doi.org/10.1128/genomeA.00115-17 |
| [38] |
Sardelic S, Bedenic B, Colinon-Dupuich C, et al. (2012) Infrequent finding of metallo-β-lactamase VIM-2 in carbapenem-resistant Pseudomonas aeruginosa strains from Croatia. Antimicrob Agents Chemother 56: 2746-2749. https://doi.org/10.1128/AAC.05212-11
|
| [39] |
Guzvinec M, Izdebski R, Butic I, et al. (2014) Sequence types 235, 111, and 132 predominate among multidrug-resistant Pseudomonas aeruginosa clinical isolates in Croatia. Antimicrob Agents Chemother 58: 6277-6283. https://doi.org/10.1128/AAC.03116-14
|
| [40] |
Krilanović M, Tomić-Paradžik M, Meštrović T, et al. (2020) Extended-spectrum β-lactamases and plasmid diversity in urinary isolates of Escherichia coli in Croatia: a nation-wide, multicentric, retrospective study. Folia Microbiol (Praha) 65: 649-667. https://doi.org/10.1007/s12223-019-00769-1
|
| [41] |
Bandić Pavlović D, Pospišil M, Nađ M, et al. (2024) Multidrug-resistant bacteria in surgical intensive care units: Antibiotic susceptibility and β-lactamase characterization. Pathogens 13: 411. https://doi.org/10.3390/pathogens13050411
|
| [42] |
D'Onofrio V, Conzemius R, Varda-Brkić D, et al. (2020) Epidemiology of colistin-resistant, carbapenemase-producing Enterobacteriaceae and Acinetobacter baumannii in Croatia. Infect Genet Evol 81: 104263. https://doi.org/10.1016/j.meegid.2020.104263
|
| [43] |
Petrović T, Uzunović S, Barišić I, et al. (2018) Arrival of carbapenem-hydrolyzing-oxacillinases in Acinetobacter baumannii in Bosnia and Herzegovina. Infect Genet Evol 58: 192-198. https://doi.org/10.1016/j.meegid.2017.12.021
|
| [44] |
Ibrahimagić A, Bedenić B, Kamberović F, et al. (2015) High prevalence of CTX-M-15 and first report of CTX-M-3, CTX-M-22, CTX-M-28 and plasmid-mediated AmpC β-lactamase producing Enterobacteriaceae causing urinary tract infections in Bosnia and Herzegovina in hospital and community settings. J Infect Chemother 21: 363-369. https://doi.org/10.1016/j.jiac.2015.01.003
|
| [45] |
Kompes G, Duvnjak S, Reil I, et al. (2023) First report and characterization of the mcr-1 positive multidrug-resistant Escherichia coli strain isolated from pigs in Croatia. Microorganisms 11: 2442. https://doi.org/10.3390/microorganisms11102442
|
| [46] |
Maravić A, Skočibušić M, Samanić I, et al. (2012) Antibiotic susceptibility profiles and first report of TEM extended-spectrum β-lactamase in Pseudomonas fluorescens from coastal waters of the Kaštela Bay, Croatia. World J Microbiol Biotechnol 28: 2039-2045. https://doi.org/10.1007/s11274-012-1006-5
|
| [47] |
Puljko A, Rozman SD, Barišić I, et al. (2023) Resistance to critically important antibiotics in hospital wastewater from the largest Croatian city. Sci Total Environ 870: 161805. https://doi.org/10.1016/j.scitotenv.2023.161805
|
| [48] |
Hrenovic J, Goic-Barisic I, Kazazic S, et al. (2016) Carbapenem-resistant isolates of Acinetobacter baumannii in a municipal wastewater treatment plant, Croatia, 2014. Euro Surveill 21. https://doi.org/10.2807/1560-7917.ES.2016.21.15.30195
|
| [49] |
Hrenovic J, Durn G, Music MS, et al. (2017) Extensively and multi drug-resistant Acinetobacter baumannii recovered from technosol at a dump site in Croatia. Sci Total Environ 607–608: 1049-1055. https://doi.org/10.1016/j.scitotenv.2017.07.108
|
| [50] |
Higgins PG, Hrenovic J, Seifert H, et al. (2018) Characterization of Acinetobacter baumannii from water and sludge line of secondary wastewater treatment plant. Water Res 140: 261-267. https://doi.org/10.1016/j.watres.2018.04.057
|
| [51] |
Cirkovic I, Muller BH, Janjusevic A, et al. (2023) Whole-Genome sequencing snapshot of clinically relevant Carbapenem-Resistant Gram-Negative bacteria from wastewater in Serbia. Antibiotics 12: 350. https://doi.org/10.3390/antibiotics12020350
|
| [52] |
Velhner M, Todorović D, Novović K, et al. (2021) Characterization of antibiotic resistance in Escherichia coli isolates from Black-headed gulls (Larus ridibundus) present in the city of Novi Sad, Serbia. Vet Res Commun 45: 199-209. https://doi.org/10.1007/s11259-021-09801-7
|
| [53] |
Libisch B, Muzslay M, Gacs M, et al. (2006) Molecular epidemiology of VIM-4 metallo-β-lactamase-producing Pseudomonas sp. isolates in Hungary. Antimicrob Agents Chemother 50: 4220-4223. https://doi.org/10.1128/AAC.00300-06
|
| [54] |
Libisch B, Watine J, Balogh B, et al. (2008) Molecular typing indicates an important role for two international clonal complexes in dissemination of VIM-producing Pseudomonas aeruginosa clinical isolates in Hungary. Res Microbiol 159: 162-168. https://doi.org/10.1016/j.resmic.2007.12.008
|
| [55] |
Szabó D, Szentandrássy J, Juhász Z, et al. (2008) Imported PER-1 producing Pseudomonas aeruginosa, PER-1 producing Acinetobacter baumannii and VIM-2-producing Pseudomonas aeruginosa strains in Hungary. Ann Clin Microbiol Antimicrob 7: 12. https://doi.org/10.1186/1476-0711-7-12
|
| [56] |
Libisch B, Balogh B, Füzi M (2009) Identification of two multidrug-resistant Pseudomonas aeruginosa clonal lineages with a countrywide distribution in Hungary. Curr Microbiol 58: 111-116. https://doi.org/.1007/s00284-008-9285-7
|
| [57] |
O'Neall D, Juhász E, Tóth Á, et al. (2020) Ceftazidime-avibactam and ceftolozane-tazobactam susceptibility of multidrug resistant Pseudomonas aeruginosa strains in Hungary. Acta Microbiol Immunol Hung 67: 61-65. https://doi.org/10.1556/030.2020.01152
|
| [58] |
Szmolka A, Anjum MF, La Ragione RM, et al. (2012) Microarray based comparative genotyping of gentamicin resistant Escherichia coli strains from food animals and humans. Vet Microbiol 156: 110-118. https://doi.org/10.1016/j.vetmic.2011.09.030
|
| [59] |
Tóth Á, Juhász-Kaszanyitzky E, Mag T, et al. (2013) Characterization of extended-spectrum β-lactamase (ESBL) producing Escherichia coli strains isolated from animal and human clinical samples in Hungary in 2006–2007. Acta Microbiol Immunol Hung 60: 175-185. https://doi.org/10.1556/AMicr.60.2013.2.8
|
| [60] |
Gulyás D, Kamotsay K, Szabó D, et al. (2023) Investigation of delafloxacin resistance in Multidrug-Resistant Escherichia coli strains and the detection of E. coli ST43 international high-risk clone. Microorganisms 11: 1602. https://doi.org/10.3390/microorganisms11061602
|
| [61] |
Nagy JB, Koleszár B, Khayer B, et al. (2023) Carbapenem-resistant Escherichia coli in Black-headed gulls, the Danube, and human clinical samples: A One Health comparison of contemporary isolates. J Glob Antimicrob Resist 35: 257-261. https://doi.org/10.1016/j.jgar.2023.10.002
|
| [62] |
Balázs B, Tóth Z, Nagy JB, et al. (2022) Faecal carriage of Carbapenem-Resistant Acinetobacter baumannii: comparison to clinical isolates from the same period (2017–2019). Pathogens 11: 1003. https://doi.org/10.3390/pathogens11091003
|
| [63] |
Szmolka A, Lestár B, Pászti J, et al. (2015) Conjugative IncF and IncI1 plasmids with tet(A) and class 1 integron conferring multidrug resistance in F18+ porcine enterotoxigenic E. coli. Acta Vet Hung 63: 425-443. https://doi.org/10.1556/004.2015.040
|
| [64] | Libisch B, Nagy G, Csivincsik A, et al. (2023) Detection of acquired antibiotic resistance determinants in the intestinal microbiota of food producing animals in Hungary. Centre of expertise on antimicrobial consumption and resistance in animals (AMCRA) (Editor) Book of Abstracts, “Antimicrobial resistance: A challenge for public health, animal health and environment” . Belgium: Scientific Symposium Brussels 19-20. |
| [65] | Libisch B, Abdulkadir S, Papp P, et al. (2022) Detection of acquired antibiotic resistance genes in domestic pig (Sus scrofa) and common carp (Cyprinus carpio) intestinal samples by metagenomics analyses in Hungary. Antibiotics 1441. https://doi.org/10.3390/antibiotics11101441 |
| [66] |
Szmolka A, Gellért Á, Szemerits D, et al. (2023) Emergence and Genomic Features of a mcr-1 Escherichia coli from Duck in Hungary. Antibiotics 12: 1519. https://doi.org/10.3390/antibiotics12101519
|
| [67] |
Libisch B, Keresztény T, Kerényi Z, et al. (2020) Metagenomic analysis of acquired antibiotic resistance determinants in the gut microbiota of wild boars (Sus Scrofa)-Preliminary Results. J Vet Res 64: 111-118. https://doi.org/10.2478/jvetres-2020-0015
|
| [68] | Hammoudi Halat D, Ayoub Moubareck C (2022) The intriguing carbapenemases of Pseudomonas aeruginosa: Current status, genetic profile, and global epidemiology. Yale J Biol Med 95: 507-515. |
| [69] | Hadadi-Fishani M, Khaledi A, Fatemi-Nasab ZS (2020) Correlation between biofilm formation and antibiotic resistance in Pseudomonas aeruginosa: A meta-analysis. Infez Med 28: 47-54. |
| [70] |
Qin S, Xiao W, Zhou C, et al. (2022) Pseudomonas aeruginosa: pathogenesis, virulence factors, antibiotic resistance, interaction with host, technology advances and emerging therapeutics. Sig Transduct Target Ther 7: 199. https://doi.org/10.1038/s41392-022-01056-1
|
| [71] |
Poirel L, Madec J, Lupo A, et al. (2018) Antimicrobial resistance in Escherichia coli. Microbiol Spectr 6. https://doi.org/10.1128/microbiolspec.arba-0026-2017
|
| [72] |
Ibrahim S, Al-Saryi N, Al-Kadmy IMS, et al. (2021) Multidrug-resistant Acinetobacter baumannii as an emerging concern in hospitals. Mol Biol Rep 48: 6987-6998. https://doi.org/10.1007/s11033-021-06690-6
|
| [73] | Mukhopadhyay H, Bairagi A, Mukherjee A, et al. (2024) Multidrug resistant Acinetobacter baumannii: A study on its pathogenesis and therapeutics. Cur Res Micro Sci 8: 100331. https://api.semanticscholar.org/CorpusID:274579668 |
| [74] | Guidelines for the prevention and control of carbapenem-resistant Enterobacteriaceae, Acinetobacter baumannii and Pseudomonas aeruginosa in health care facilities. Available from: https://iris.who.int/bitstream/handle/10665/259462/9789241550178-eng.pdf |
| [75] |
Yong D, Toleman MA, Giske CG, et al. (2009) Characterization of a new metallo-β-lactamase gene, blaNDM-1, and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. AAC 53: 5046-5054. https://doi.org/10.1128/AAC.00774-09
|
| [76] |
Kehl K, Schallenberg A, Szekat C, et al. (2022) Dissemination of carbapenem resistant bacteria from hospital wastewater into the environment. Sci Total Environ 806: 151339. https://doi.org/10.1016/j.scitotenv.2021.151339
|
| [77] |
Zagui GS, Gonçalves de Almeida OG, Moreira NC (2024) Hospital wastewater as source of human pathogenic bacteria: A phenotypic and genomic analysis of international high-risk clone VIM-2-producing Pseudomonas aeruginosa ST235/O11. Environ Res 255: 119166. https://doi.org/10.1016/j.envres.2024.119166
|
| [78] |
Jangsangthong A, Lugsomya K, Apiratwarrasakul S, et al. (2024) Distribution of sequence types and antimicrobial resistance of clinical Pseudomonas aeruginosa isolates from dogs and cats visiting a veterinary teaching hospital in Thailand. BMC Vet Res 20: 234. https://doi.org/10.1186/s12917-024-04098-5
|
| [79] | Hayashi W, Izumi K, Yoshida S, et al. (2021) Antimicrobial resistance and Type III secretion system virulotypes of Pseudomonas aeruginosa isolates from dogs and cats in Primary Veterinary Hospitals in Japan: Identification of the international high-risk clone sequence type 235. Microbiol Spectr 9. https://doi.org/10.1128/Spectrum.00408-21 |
| [80] | Soliman AM, Nariya H, Tanaka D, et al. (2021) Vegetable-derived carbapenemase-producing high-risk Klebsiella pneumoniae ST15 and Acinetobacter baumannii ST2 clones in Japan: Coexistence of blaNDM-1, blaOXA-66, blaOXA-72, and an AbaR4-Like resistance island in the same sample. Appl Environ Microbiol 8. https://doi.org/10.1128/AEM.02166-20 |
| [81] | Taj Z, Rasool MH, Almatroudi A, et al. (2020) Extensively Drug-resistant Acinetobacter baumannii belonging to international clone II from a pet cat with urinary tract Infection; The first report from Pakistan. Pol J Microbiol 69: 1-4. https://doi.org/10.33073/pjm-2020-017 |
| [82] |
Mshana SE, Imirzalioglu C, Hain T, et al. (2011) Multiple ST clonal complexes, with a predominance of ST131, of Escherichia coli harbouring blaCTX-M-15 in a tertiary hospital in Tanzania. Clin Microbiol Infect 17: 1279-1282. https://doi.org/10.1111/j.1469-0691.2011.03518.x
|
| [83] |
Jouini A, Klibi A, Elarbi I, et al. (2021) First detection of human ST131-CTX-M-15-O25-B2 Clone and High-Risk clonal lineages of ESBL/pAmpC-Producing E. coli isolates from diarrheic poultry in Tunisia. Antibiotics 10: 670. https://doi.org/10.3390/antibiotics10060670
|
| [84] |
Hernandez J, Bonnedahl J, Eliasson I, et al. (2010) Globally disseminated human pathogenic Escherichia coli of O25b-ST131 clone, harbouring blaCTX-M-15, found in Glaucous-winged gull at remote Commander Islands, Russia. Environ Microbiol Rep 2: 329-332. https://doi.org/10.1111/j.1758-2229.2010.00142.x
|
| [85] |
Pandey S, Doo H, Keum GB, et al. (2023) Antibiotic resistance in livestock, environment and humans: One Health perspective. Anim Sci Technol 66: 266-278. https://doi.org/10.5187/jast.2023.e129
|
| [86] |
Libisch B (2024) N-Alkane Assimilation by Pseudomonas aeruginosa and its interactions with virulence and antibiotic resistance. Antibiotics 13: 1028. https://doi.org/10.3390/antibiotics13111028
|
| [87] |
Libisch B, Picot C, Ceballos-Garzon A, et al. (2022) Prototheca infections and ecology from a One Health perspective. Microorganisms 10: 938. https://doi.org/10.3390/microorganisms10050938
|
| [88] |
Martak D, Henriot CP, Didier Hocquet D (2024) Environment, animals, and food as reservoirs of antibiotic-resistant bacteria for humans: One health or more?. Infectious Diseases Now 54: 104895. https://doi.org/10.1016/j.idnow.2024.104895
|
| [89] |
Ozoaduche CL, Libisch B, Itoro D, et al. (2025) Antibiotic resistance and virulence determinants of Pseudomonas aeruginosa isolates cultured from Hydrocarbon-Contaminated environmental samples. Microorganisms 13: 688. https://doi.org/10.3390/microorganisms13030688
|
| [90] |
Zhou N, Cheng Z, Zhang X, et al. (2022) Global antimicrobial resistance: A system-wide comprehensive investigation using the Global One Health Index. Infect Dis Poverty 11: 92. https://doi.org/10.1186/s40249-022-01016-5
|
| [91] | GBD 2021 Antimicrobial Resistance Collaborators.Global burden of bacterial antimicrobial resistance 1990–2021: A systematic analysis with forecasts to 2050. Lancet (2024) 404: 1199-1226. https://doi.org/10.1016/S0140-6736(24)01867-1 |
 microbiol-11-02-020-s001.xlsx microbiol-11-02-020-s001.xlsx |
 |
Figures(2) / Tables(7)

Chioma Lilian Ozoaduche, Katalin Posta, Balázs Libisch, Ferenc Olasz. Acquired antibiotic resistance of Pseudomonas spp., Escherichia coli and Acinetobacter spp. in the Western Balkans and Hungary with a One Health outlook[J]. AIMS Microbiology, 2025, 11(2): 436-461. doi: 10.3934/microbiol.2025020







 DownLoad:
DownLoad: