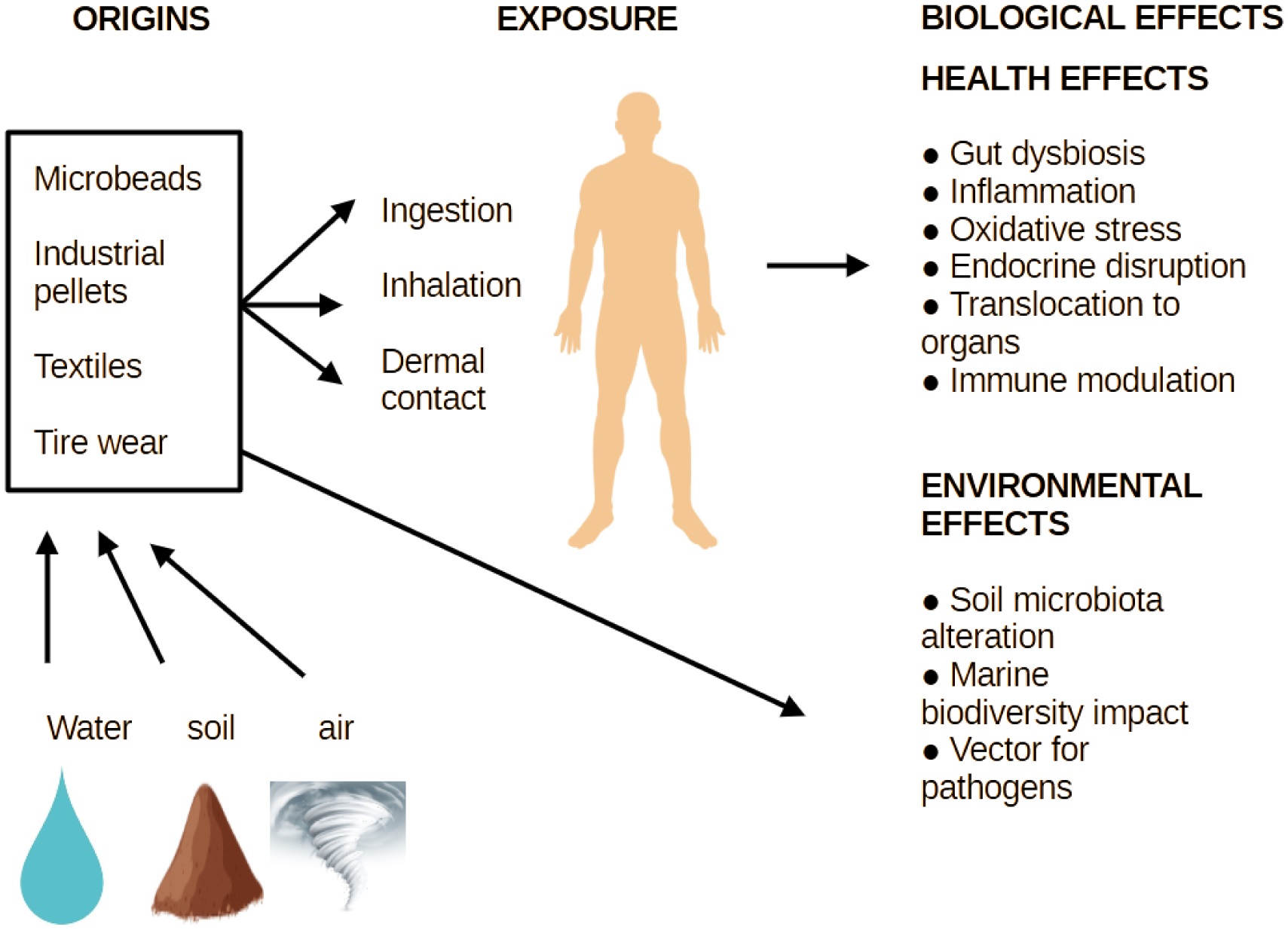
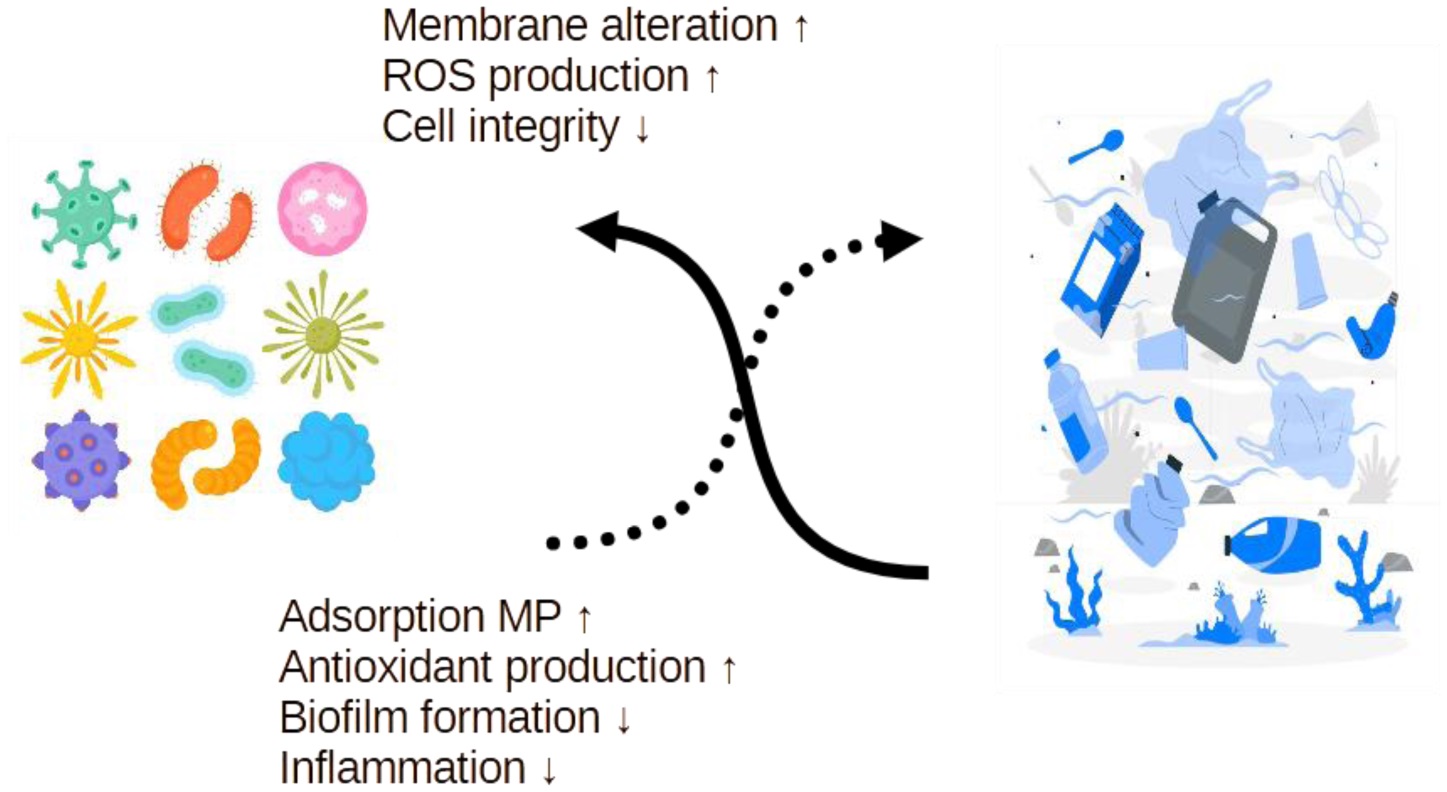
Microplastics (MPs), synthetic polymer particles less than 5 mm in size, are an emerging contaminant with implications for both human and ecosystem health. Being widespread in food and water sources, MPs can disrupt gastrointestinal integrity, alter the microbiota composition, and provoke oxidative and inflammatory responses. Probiotics, live microorganisms known for their gut health benefits, are now being explored for their ability to mitigate these effects. This review synthesizes evidence from in vitro and in vivo studies on how MPs impact probiotic viability, adhesion, and biofilm formation, and how certain strains may counter MP-induced toxicity by modulating oxidative stress, immune function, and the epithelial barrier integrity. Additionally, this manuscript discusses emerging applications in environmental microbiology, such as the potential use of native and engineered probiotics for microplastic bioremediation. Although the current data highlight promising avenues, key gaps remain in our understanding of strain-specific mechanisms, long-term efficacy, and real-world applicability. Addressing these will be essential to advance probiotic-based strategies in both human and environmental contexts.
Citation: Jean DEMARQUOY. Microplastics and probiotics: Mechanisms of interaction and their consequences for health[J]. AIMS Microbiology, 2025, 11(2): 388-409. doi: 10.3934/microbiol.2025018
Microplastics (MPs), synthetic polymer particles less than 5 mm in size, are an emerging contaminant with implications for both human and ecosystem health. Being widespread in food and water sources, MPs can disrupt gastrointestinal integrity, alter the microbiota composition, and provoke oxidative and inflammatory responses. Probiotics, live microorganisms known for their gut health benefits, are now being explored for their ability to mitigate these effects. This review synthesizes evidence from in vitro and in vivo studies on how MPs impact probiotic viability, adhesion, and biofilm formation, and how certain strains may counter MP-induced toxicity by modulating oxidative stress, immune function, and the epithelial barrier integrity. Additionally, this manuscript discusses emerging applications in environmental microbiology, such as the potential use of native and engineered probiotics for microplastic bioremediation. Although the current data highlight promising avenues, key gaps remain in our understanding of strain-specific mechanisms, long-term efficacy, and real-world applicability. Addressing these will be essential to advance probiotic-based strategies in both human and environmental contexts.

| [1] |
Ahmad MF, Fauzi MA, Ahmad MH, et al. (2024) Are we eating plastic? Science mapping of microplastic pollution in the aquatic food chain. Integr Environ Assess Manag 20: 1800-1811. https://doi.org/10.1002/ieam.4930 
|
| [2] |
Barceló D, Picó Y, Alfarhan AH (2023) Microplastics: Detection in human samples, cell line studies, and health impacts. Environ Toxicol Pharmacol 101: 104204. https://doi.org/10.1016/j.etap.2023.104204
|
| [3] |
Zeb A, Liu W, Ali N, et al. (2024) Microplastic pollution in terrestrial ecosystems: Global implications and sustainable solutions. J Hazard Mater 461: 132636. https://doi.org/10.1016/j.jhazmat.2023.132636
|
| [4] |
Sharma S, Bhardwaj A, Thakur M, et al. (2024) Understanding microplastic pollution of marine ecosystem: a review. Environ Sci Pollut Res Int 31: 41402-14145. https://doi.org/10.1007/s11356-023-28314-1
|
| [5] |
Fröhlich E (2024) Local and systemic effects of microplastic particles through cell damage, release of chemicals and drugs, dysbiosis, and interference with the absorption of nutrients. J Toxicol Environ Health B Crit Rev 27: 315-344. https://doi.org/10.1080/10937404.2024.2406192
|
| [6] | WHO/FAOProbiotics in food, health and nutritional properties and guidelines for evaluation. Report of a joint FAO/WHO working group on drafting guidelines for the avaluation of probiotics in food (2002). |
| [7] |
Sepehr A, Miri ST, Aghamohammad S, et al. (2024) Health benefits, antimicrobial activities, and potential applications of probiotics: A review. Medicine (Baltimore) 103: e32412. https://doi/10.1097/MD.0000000000032412
|
| [8] |
Lázaro Á, Vila-Donat P, Manyes L (2024) Emerging mycotoxins and preventive strategies related to gut microbiota changes: probiotics, prebiotics, and postbiotics-a systematic review. Food Funct 15: 8998-9023. https://doi.org/10.1039/D4FO01705F
|
| [9] |
Ghobish SA, Motti CA, Bissember AC, et al. (2025) Microplastics in the marine environment: Challenges and the shift towards sustainable plastics and plasticizers. J Hazard Mater 491: 137945. https://doi.org/10.1016/j.jhazmat.2025.137945
|
| [10] |
Aransiola SA, Victor-Ekwebelem MO, Daza BX, et al. (2025) Micro- and nano-plastics pollution in the marine environment: Progresses, drawbacks and future guidelines. Chemosphere 374: 144211. https://doi.org/10.1016/j.chemosphere.2025.144211
|
| [11] |
Frias JPGL, Nash R (2019) Microplastics: Finding a consensus on the definition. Mar Pollut Bull 138: 145-147. https://doi.org/10.1016/j.marpolbul.2018.11.022
|
| [12] |
Browne MA, Crump P, Niven SJ, et al. (2011) Accumulation of microplastic on shorelines woldwide: sources and sinks. Environ Sci Technol 45: 9175-9179. https://doi.org/10.1016/j.marpolbul.2018.11.022
|
| [13] |
Wright SL, Kelly FJ (2017) Plastic and human health: A micro issue?. Environ Sci Technol 51: 6634-6647. https://doi.org/10.3390/metabo12030223
|
| [14] |
Schwarzfischer M, Rogler G (2022) The intestinal barrier-shielding the body from nano- and microparticles in our diet. Metabolites 12: 223. https://doi.org/10.3390/metabo12030223
|
| [15] |
Tanoiri H, Nakano H, Arakawa H, et al. (2021) Inclusion of shape parameters increases the accuracy of 3D models for microplastics mass quantification. Mar Pollut Bull 171: 112749. https://doi.org/10.1016/j.marpolbul.2021.112749
|
| [16] |
Schlundt C, Mark Welch JL, Knochel AM, et al. (2020) Spatial structure in the “Plastisphere”: Molecular resources for imaging microscopic communities on plastic marine debris. Mol Ecol Resour 20: 620-634. https://doi.org/10.1111/1755-0998.13119
|
| [17] |
Campanale C, Massarelli C, Savino I, et al. (2020) A detailed review study on potential effects of microplastics and additives of concern on human health. Int J Environ Res Public Health 17. https://doi.org/10.3390/ijerph17041212
|
| [18] |
Rochman CM, Hoh E, Kurobe T, et al. (2013) Ingested plastic transfers hazardous chemicals to fish and induces hepatic stress. Sci Rep 3: 3263. https://doi.org/10.1038/srep03263
|
| [19] |
Taglialegna A (2023) Microplastics impair seabird gut microbiomes. Nat Rev Microbiol 21: 343. https://doi.org/10.1038/s41579-023-00893-3
|
| [20] | Yong CQY, Valiyaveettil S, Tang BL (2020) Toxicity of microplastics and nanoplastics in mammalian systems. Int J Environ Res Public Health 17. https://doi.org/10.3390/ijerph17051509 |
| [21] |
Bocker R, Silva EK (2025) Microplastics in our diet: A growing concern for human health. Sci Total Environ 968: 178882. https://doi.org/10.1016/j.scitotenv.2025.178882
|
| [22] |
Hartmann NB, Hüffer T, Thompson RC, et al. (2019) Are we speaking the same language? recommendations for a definition and categorization framework for plastic debris. Environ Sci Technol 53: 1039-1047. https://doi.org/10.1021/acs.est.8b05297
|
| [23] |
Prabhu K, Ghosh S, Sethulekshmi S, et al. (2024) In vitro digestion of microplastics in human digestive system: Insights into particle morphological changes and chemical leaching. Sci Total Environ 934: 173173. https://doi.org/10.1016/j.scitotenv.2024.173173
|
| [24] |
Chandra S, Walsh KB (2024) Microplastics in water: Occurrence, fate and removal. J Contam Hydrol 264: 104360. https://doi.org/10.1016/j.jconhyd.2024.104360
|
| [25] |
Yang L, Kang S, Luo X, et al. (2024) Microplastics in drinking water: A review on methods, occurrence, sources, and potential risks assessment. Environ Pollut 348: 123857.
|
| [26] |
Nicole W (2021) Microplastics in seafood: How much are people eating?. Environ Health Perspect 129: 34001. https://doi.org/10.1016/j.envpol.2024.123857
|
| [27] |
Zhang Q, Xu EG, Li J, et al. (2020) A review of microplastics in table salt, drinking water, and air: Direct human exposure. Environ Sci Technol 54: 3740-3751. https://doi.org/10.1021/acs.est.9b04535
|
| [28] |
Alma AM, de Groot GS, Buteler M (2023) Microplastics incorporated by honeybees from food are transferred to honey, wax and larvae. Environ Pollut 320: 121078. https://doi.org/10.1016/j.envpol.2023.121078
|
| [29] |
Kaseke T, Lujic T, Cirkovic Velickovic T (2023) Nano- and microplastics migration from plastic food packaging into dairy products: impact on nutrient digestion, absorption, and metabolism. Foods 12. https://doi.org/10.3390/foods12163043
|
| [30] |
Cox KD, Covernton GA, Davies HL, et al. (2019) Human consumption of microplastics. Environ Sci Technol 53: 7068-7074. https://doi.org/10.1021/acs.est.9b01517
|
| [31] |
Gaspar L, Bartman S, Coppotelli G, et al. (2023) Acute exposure to microplastics induced changes in behavior and inflammation in young and old mice. Int J Mol Sci 24. https://doi.org/10.3390/ijms241512308
|
| [32] |
Jin Y, Lu L, Tu W, et al. (2019) Impacts of polystyrene microplastic on the gut barrier, microbiota and metabolism of mice. Sci Total Environ 649: 308-317. https://doi.org/10.1016/j.scitotenv.2018.08.353
|
| [33] |
Lu L, Wan Z, Luo T, et al. (2018) Polystyrene microplastics induce gut microbiota dysbiosis and hepatic lipid metabolism disorder in mice. Sci Total Environ 631–632: 449-458. https://doi.org/10.1016/j.scitotenv.2018.03.051
|
| [34] |
Deng Y, Zhang Y, Lemos B, et al. (2017) Tissue accumulation of microplastics in mice and biomarker responses suggest widespread health risks of exposure. Sci Rep 7: 46687. https://doi.org/10.1038/srep46687
|
| [35] |
Li B, Ding Y, Cheng X, et al. (2020) Polyethylene microplastics affect the distribution of gut microbiota and inflammation development in mice. Chemosphere 244: 125492. https://doi.org/10.1016/j.chemosphere.2019.125492
|
| [36] |
Bao X, Gu Y, Chen L, et al. (2024) Microplastics derived from plastic mulch films and their carrier function effect on the environmental risk of pesticides. Sci Total Environ 924: 171472. https://doi.org/10.1016/j.scitotenv.2024.171472
|
| [37] |
Stenger KS, Wikmark OG, Bezuidenhout CC, et al. (2021) Microplastics pollution in the ocean: Potential carrier of resistant bacteria and resistance genes. Environ Pollut 291: 118130. https://doi.org/10.1016/j.envpol.2021.118130
|
| [38] |
Demarquoy J (2024) Microplastics and microbiota: Unraveling the hidden environmental challenge. World J Gastroenterol 30: 2191-2194. https://doi.org/10.3748/wjg.v30.i16.2191
|
| [39] |
Giri SS, Kim HJ, Jung WJ, et al. (2024) Probiotics in addressing heavy metal toxicities in fish farming: Current progress and perspective. Ecotoxicol Environ Saf 282: 116755. https://doi.org/10.1016/j.ecoenv.2024.116755
|
| [40] |
Emanowicz P, Średnicka P, Wójcicki M, et al. (2024) Mitigating dietary bisphenol exposure through the gut microbiota: The role of next-generation probiotics in bacterial detoxification. Nutrients 16. https://doi.org/10.3390/nu16213757
|
| [41] |
Bazeli J, Banikazemi Z, Hamblin MR, et al. (2023) Could probiotics protect against human toxicity caused by polystyrene nanoplastics and microplastics?. Front Nutr 10: 1186724. https://doi.org/10.3389/fnut.2023.1186724
|
| [42] |
Teng X, Zhang T, Rao C (2024) Novel probiotics adsorbing and excreting microplastics. Front Microbiol 15: 1522794. https://doi.org/10.3389/fmicb.2024.1522794
|
| [43] |
Chi J, Patterson JS, Jin Y, et al. (2025) Metabolic reprogramming in gut microbiota exposed to polystyrene microplastics. Biomedicines 13: 446. https://doi.org/10.3390/biomedicines13020446
|
| [44] |
Shi L, Wu C, Wang Y, et al. (2025) Lactobacillus plantarum reduces polystyrene microplastic induced toxicity via multiple pathways: A potentially effective and safe dietary strategy to counteract microplastic harm. J Hazard Mater 489: 137669. https://doi.org/10.1016/j.envpol.2024.125288
|
| [45] |
Athulya PA, Chandrasekaran N, Thomas J (2024) Polystyrene microplastics interaction and influence on the growth kinetics and metabolism of tilapia gut probiotic. Environ Sci Process Impacts 26: 221-232. https://doi.org/10.1039/d3em00369h
|
| [46] |
Salas-Jara MJ, Ilabaca A, Vega M, et al. (2016) Biofilm forming lactobacillus: New challenges for the development of probiotics. Microorganisms 4: 35. https://doi.org/10.3390/microorganisms4030035
|
| [47] |
Sharma S, Kanwar SS (2017) Adherence potential of indigenous lactic acid bacterial isolates obtained from fermented foods of Western Himalayas to intestinal epithelial Caco-2 and HT-29 cell lines. J Food Sci Technol 54: 3504-3511. https://doi.org/10.1007/s13197-017-2807-1
|
| [48] |
Liu S, Junaid M, Liao H, et al. (2022) Eco-corona formation and associated ecotoxicological impacts of nanoplastics in the environment. Sci Total Environ 836: 155703. https://doi.org/10.1016/j.scitotenv.2022.155703
|
| [49] |
Fackelmann G, Sommer S (2019) Microplastics and the gut microbiome: How chronically exposed species may suffer from gut dysbiosis. Mar Pollut Bull 143: 193-203. https://doi.org/10.1016/j.marpolbul.2019.04.030
|
| [50] |
Jia R, Han J, Liu X, et al. (2023) Exposure to polypropylene microplastics via oral ingestion induces colonic apoptosis and intestinal barrier damage through oxidative stress and inflammation in Mice. Toxics 11: 127. https://doi.org/10.3390/toxics11020127
|
| [51] |
Jia R, Han J, Liu X, et al. (2023) Correction: Jia et al. Exposure to polypropylene microplastics via oral ingestion induces colonic apoptosis and intestinal barrier damage through oxidative stress and inflammation in Mice. Toxics 2023, 11, 127. Toxics 11: 733. https://doi.org/10.3390/toxics11090733
|
| [52] |
Sincihu Y, Lusno MFD, Mulyasari TM, et al. (2023) Wistar rats hippocampal neurons response to blood low-density polyethylene microplastics: A pathway analysis of SOD, CAT, MDA, 8-OHdG expression in hippocampal neurons and blood serum Aβ42 levels. Neuropsychiatr Dis Treat 19: 73-83. https://doi.org/10.2147/NDT.S396556
|
| [53] |
Zhang Y, Hou B, Liu T, et al. (2023) Probiotics improve polystyrene microplastics-induced male reproductive toxicity in mice by alleviating inflammatory response. Ecotoxicol Environ Saf 263: 115248. https://doi.org/10.1016/j.ecoenv.2023.115248
|
| [54] |
Fleury JB, Baulin VA (2021) Microplastics destabilize lipid membranes by mechanical stretching. Proc Natl Acad Sci U S A 118. https://doi.org/10.1073/pnas.2104610118
|
| [55] |
Järvenpää J, Perkkiö M, Laitinen R, et al. (2022) PE and PET oligomers' interplay with membrane bilayers. Sci Rep 12: 2234. https://doi.org/10.1038/s41598-022-06217-4
|
| [56] |
Kadac-Czapska K, Ośko J, Knez E, et al. (2024) Microplastics and oxidative stress-current problems and prospects. Antioxidants (Basel) 13: 579. https://doi.org/10.3390/antiox13050579
|
| [57] |
Wu Z, Li Y, Chen H, et al. (2025) Investigation of microplastics from granulated virgin and UV aged samples in artificial digestion system. Polymer Degrad Stab 232: 111157. https://doi.org/10.1016/j.polymdegradstab.2024.111157
|
| [58] |
Li H, Xu S, Zhou F, et al. (2024) Polystyrene microplastics exposure: Disruption of intestinal barrier integrity and hepatic function in infant mice. Ecotoxicol Environ Saf 288: 117357. https://doi.org/10.1016/j.ecoenv.2024.117357
|
| [59] |
Chen PY, Li S, Koh YC, et al. (2019) Oolong tea extract and citrus peel polymethoxyflavones reduce transformation of l-carnitine to trimethylamine-. J Agric Food Chem 67: 7869-7879. https://doi.org/10.1021/acs.jafc.9b03092
|
| [60] |
Bora SS, Gogoi R, Sharma MR, et al. (2024) Microplastics and human health: unveiling the gut microbiome disruption and chronic disease risks. Front Cell Infect Microbiol 14: 1492759. https://doi.org/10.3389/fcimb.2024.1492759
|
| [61] |
Zhao L, Dou Q, Chen S, et al. (2023) Adsorption abilities and mechanisms of Lactobacillus on various nanoplastics. Chemosphere 320: 138038. https://doi.org/10.1016/j.chemosphere.2023.138038
|
| [62] |
Rafa N, Ahmed B, Zohora F, et al. (2024) Microplastics as carriers of toxic pollutants: Source, transport, and toxicological effects. Environ Pollut 343: 123190. https://doi.org/10.1016/j.envpol.2023.123190
|
| [63] |
Chen H, Nan S, Liu H, et al. (2022) Investigation of microplastics in digestion system: effect on surface microstructures and probiotics. Bull Environ Contam Toxicol 109: 882-892. https://doi.org/10.1007/s00128-022-03571-x
|
| [64] |
Gao J, Sadiq FA, Zheng Y, et al. (2022) Biofilm-based delivery approaches and specific enrichment strategies of probiotics in the human gut. Gut Microbes 14: 2126274. https://doi.org/10.1080/19490976.2022.2126274
|
| [65] |
Kim JH, Ruegger PR, Lebig EG, et al. (2020) High levels of oxidative stress create a microenvironment that significantly decreases the diversity of the microbiota in diabetic chronic wounds and promotes biofilm formation. Front Cell Infect Microbiol 10: 259. https://doi.org/10.3389/fcimb.2020.00259
|
| [66] |
Gou HZ, Zhang YL, Ren LF, et al. (2022) How do intestinal probiotics restore the intestinal barrier?. Front Microbiol 13: 929346. https://doi.org/10.3389/fmicb.2022.929346
|
| [67] |
Guo YT, Peng YC, Yen HY, et al. (2022) Effects of probiotic supplementation on immune and inflammatory markers in athletes: A meta-analysis of randomized clinical trials. Medicina (Kaunas) 58: 1188. https://doi.org/10.3390/medicina58091188
|
| [68] | Wang K, Dong H, Qi Y, et al. (2017) Lactobacillus casei regulates differentiation of Th17/Treg cells to reduce intestinal inflammation in mice. Can J Vet Res 81: 122-128. |
| [69] |
Nie X, Li Q, Ji H, et al. (2025) Bifidobacterium longum NSP001-derived extracellular vesicles ameliorate ulcerative colitis by modulating T cell responses in gut microbiota-(in)dependent manners. NPJ Biofilms Microbiomes 11: 27. https://doi.org/10.1038/s41522-025-00663-4
|
| [70] | Wu S, Wei F, Chen Y, et al. (2025) Lactiplantibacillus plantarum ZJ316 alleviates Helicobacter pylori-induced intestinal inflammation by sustaining intestinal homeostasis. Probiotics Antimicrob Proteins . https://doi.org/10.1007/s12602-025-10474-w |
| [71] |
Vermeulen R, Schymanski EL, Barabási AL, et al. (2020) The exposome and health: Where chemistry meets biology. Science 367: 392-396. https://doi.org/10.1126/science.aay3164
|
| [72] |
Barboza LGA, Vieira LR, Branco V, et al. (2018) Microplastics cause neurotoxicity, oxidative damage and energy-related changes and interact with the bioaccumulation of mercury in the European seabass, Dicentrarchus labrax (Linnaeus, 1758). Aquat Toxicol 195: 49-57. https://doi.org/10.1016/j.aquatox.2017.12.008
|
| [73] |
Prata JC, da Costa JP, Lopes I, et al. (2020) Environmental exposure to microplastics: An overview on possible human health effects. Sci Total Environ 702: 134455. https://doi.org/10.1016/j.scitotenv.2019.134455
|
| [74] |
Li D, Shi Y, Yang L, et al. (2020) Microplastic release from the degradation of polypropylene feeding bottles during infant formula preparation. Nat Food 1: 746-754. https://doi.org/10.1038/s43016-020-00171-y
|
| [75] |
Medley EA, Spratlen MJ, Yan B, et al. (2023) A systematic review of the placental translocation of micro- and nanoplastics. Curr Environ Health Rep 10: 99-111. https://doi.org/10.1007/s40572-023-00391-x
|
| [76] |
Ragusa A, Svelato A, Santacroce C, et al. (2021) Plasticenta: First evidence of microplastics in human placenta. Environ Int 146: 106274. https://doi.org/10.1016/j.envint.2020.106274
|
| [77] | Chen WC, Quigley EM (2014) Probiotics, prebiotics & synbiotics in small intestinal bacterial overgrowth: opening up a new therapeutic horizon!. Indian J Med Res 140: 582-584. |
| [78] |
Gouin T, Ellis-Hutchings R, Pemberton M, et al. (2024) Addressing the relevance of polystyrene nano- and microplastic particles used to support exposure, toxicity and risk assessment: implications and recommendations. Part Fibre Toxicol 21: 39. https://doi.org/10.1186/s12989-024-00599-1
|
| [79] |
Makkar H, Sriram G (2025) Advances in modeling periodontal host-microbe interactions: insights from organotypic and organ-on-chip systems. Lab Chip 25: 1342-1371. https://doi.org/10.1039/D4LC00871E
|
| [80] |
Urbanek AK, Kosiorowska KE, Mirończuk AM (2021) Current knowledge on polyethylene terephthalate degradation by genetically modified microorganisms. Front Bioeng Biotechnol 9: 771133. https://doi.org/10.3389/fbioe.2021.771133
|
| [81] |
Hossain S, Shukri ZNA, Waiho K, et al. (2024) Biodegradation of polyethylene (PE), polypropylene (PP), and polystyrene (PS) microplastics by floc-forming bacteria, Bacillus cereus strain SHBF2, isolated from a commercial aquafarm. Environ Sci Pollut Res 31: 32225-32245. https://doi.org/10.1007/s11356-024-33337-3
|
Figures(2) / Tables(1)

Jean DEMARQUOY. Microplastics and probiotics: Mechanisms of interaction and their consequences for health[J]. AIMS Microbiology, 2025, 11(2): 388-409. doi: 10.3934/microbiol.2025018







 DownLoad:
DownLoad: