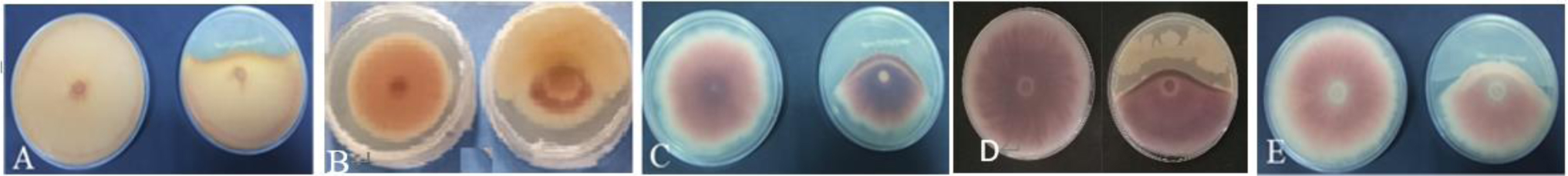
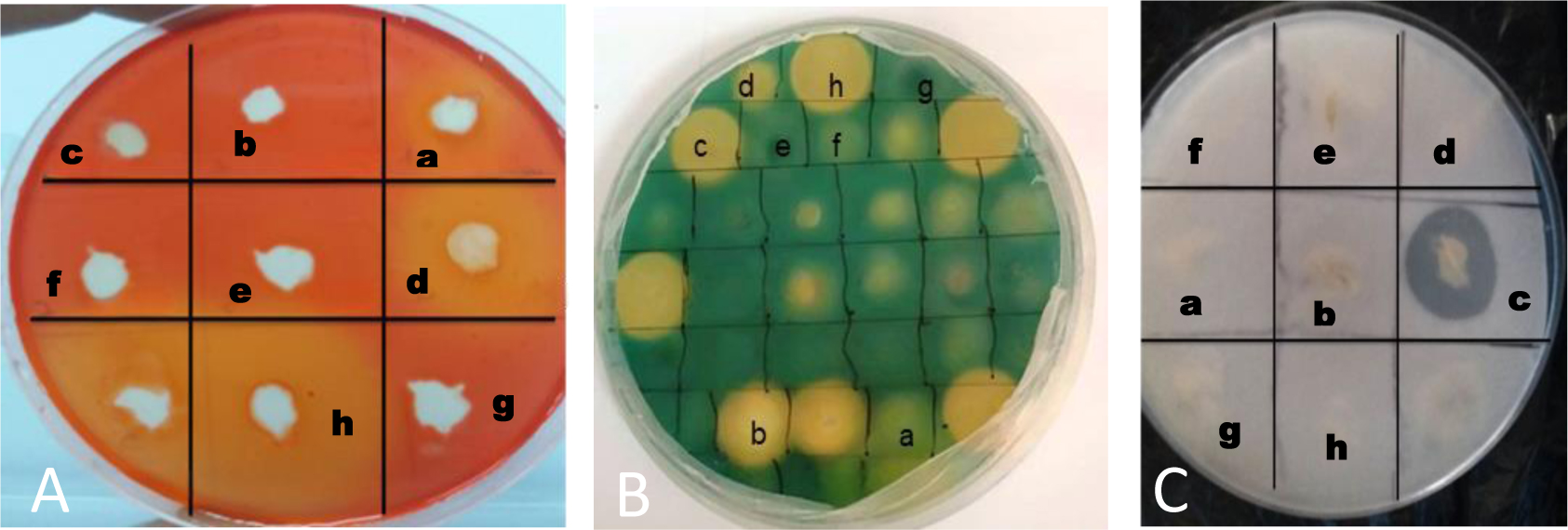
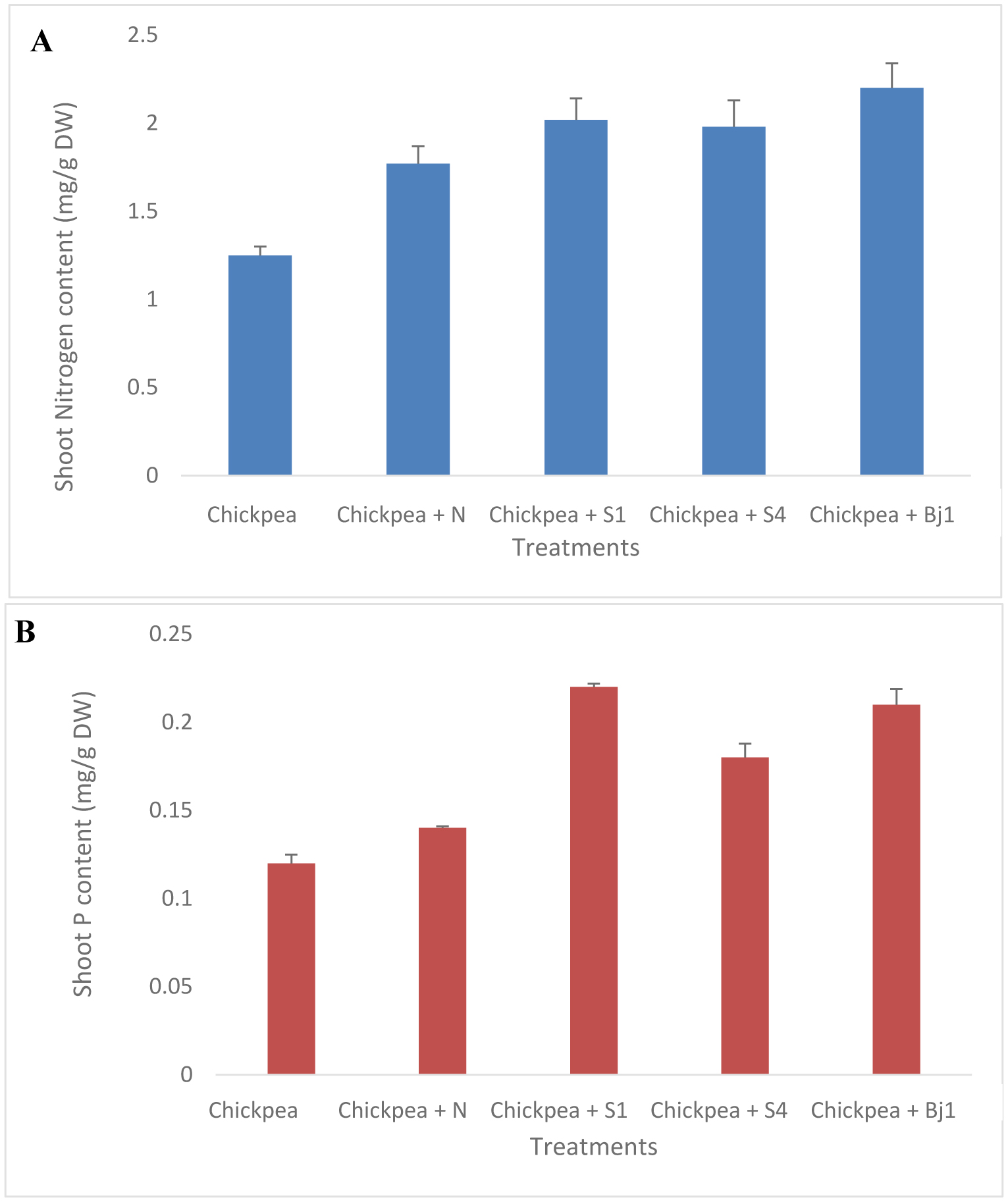
Chickpea (Cicer arietinum L.) is considered a cheap source of plant protein. In Mediterranean regions, and particularly in Tunisia, fungal attacks are likely to further aggravate drought stress and increase the economic vulnerability of chickpea production. Plant growth-promoting rhizobacteria (PGPR) and rhizobia have the potential to enhance plant growth and mitigate the adverse effects of biotic and abiotic stresses. The objective of this study was to isolate non-rhizobial rhizosphere bacteria from the soil and evaluate their ability to enhance plants' growth and symbiotic performance and to control chickpea wilt caused by F. redolens. A total of 26 bacterial isolates from rhizosphere soil samples were subsequently evaluated for their antagonistic properties against five phytopathogenic fungi (Fusarium oxysporum solani, Fusarium oxysporum matthioli, Fusarium oxysporum MN-2, Fusarium oxysporum 184, and Fusarium rdolens). Seven bacterial isolates demonstrated in vitro plant-beneficial characteristics and/or antagonistic activity against 5 Fusarium strains. Two bacterial strains including Streptomyces diastaticus subsp. diastaticus and Bacillus subtilis were chosen for additional investigation because they showed the greatest number of plant growth-promoting (PGP) traits and exhibited an antagonistic effect on pathogens. Assays conducted in pots showed that PGPRs co-inoculated with Mesorhizobium sp. Bj1 protected chickpea plants from F. redolens infection and enhanced plant growth and nutrient uptake. Pot experiments carried out in a greenhouse further demonstrated that the co-inoculation of chickpea plants with the bacterial strains and a Mesorhizobium strain lessened the severity of the F. redolens infection. These results suggest that co-inoculation with S. diastaticus subsp. diastaticus and Mesorhizobium sp. Bj1 may act as a helpful bioformulation to boost chickpea plants' growth and protect them from wilting. Other PGPR candidates included Mesorhizobium spp. and B. subtilis strains. Both Mesorhizobium sp. Bj1 and the uninoculated plants were used as controls. The association of PGPR with other inoculants potentially could substitute for chemical fertilizers, and testing of PGPR under field conditions will further elucidate their effectiveness on grain yields of chickpea.
Citation: Sabrine Balti, Yassine Mabrouk, Mouna Souihi, Imen Hemissi, Ismail Amri, Ethan Humm, Noor Khan, Ann M. Hirsch. Combined inoculation of rhizobacteria with Mesorhizobium promotes growth, nutrient contents, and protects chickpea against Fusarium redolens[J]. AIMS Microbiology, 2025, 11(2): 318-337. doi: 10.3934/microbiol.2025015
Chickpea (Cicer arietinum L.) is considered a cheap source of plant protein. In Mediterranean regions, and particularly in Tunisia, fungal attacks are likely to further aggravate drought stress and increase the economic vulnerability of chickpea production. Plant growth-promoting rhizobacteria (PGPR) and rhizobia have the potential to enhance plant growth and mitigate the adverse effects of biotic and abiotic stresses. The objective of this study was to isolate non-rhizobial rhizosphere bacteria from the soil and evaluate their ability to enhance plants' growth and symbiotic performance and to control chickpea wilt caused by F. redolens. A total of 26 bacterial isolates from rhizosphere soil samples were subsequently evaluated for their antagonistic properties against five phytopathogenic fungi (Fusarium oxysporum solani, Fusarium oxysporum matthioli, Fusarium oxysporum MN-2, Fusarium oxysporum 184, and Fusarium rdolens). Seven bacterial isolates demonstrated in vitro plant-beneficial characteristics and/or antagonistic activity against 5 Fusarium strains. Two bacterial strains including Streptomyces diastaticus subsp. diastaticus and Bacillus subtilis were chosen for additional investigation because they showed the greatest number of plant growth-promoting (PGP) traits and exhibited an antagonistic effect on pathogens. Assays conducted in pots showed that PGPRs co-inoculated with Mesorhizobium sp. Bj1 protected chickpea plants from F. redolens infection and enhanced plant growth and nutrient uptake. Pot experiments carried out in a greenhouse further demonstrated that the co-inoculation of chickpea plants with the bacterial strains and a Mesorhizobium strain lessened the severity of the F. redolens infection. These results suggest that co-inoculation with S. diastaticus subsp. diastaticus and Mesorhizobium sp. Bj1 may act as a helpful bioformulation to boost chickpea plants' growth and protect them from wilting. Other PGPR candidates included Mesorhizobium spp. and B. subtilis strains. Both Mesorhizobium sp. Bj1 and the uninoculated plants were used as controls. The association of PGPR with other inoculants potentially could substitute for chemical fertilizers, and testing of PGPR under field conditions will further elucidate their effectiveness on grain yields of chickpea.

| [1] |
Jendoubi W, Bouhadida M, Boukteb A, et al. (2017) Fusarium wilt affecting chickpea crop. Agriculture 7: 3-23. https://doi.org/10.3390/agriculture7030023 
|
| [2] | Stanisław M, Nawrocki J, Kućmierz J (2004) Disease symptoms on chickpea (Cicer arietinum L.) and their causal agents. Folia Hortic Annals 16: 47-53. |
| [3] |
Bouhadida M, Jendoubi W, Gargouri S, et al. (2017) First report of Fusarium redolens causing Fusarium yellowing and wilt of chickpea in Tunisia. Plant Dis 10: 1038-1038. https://doi.org/10.1094/PDIS-08-16-1114-PDN
|
| [4] |
Jimenez-Fernandez D, Navas-Cortes JA, Montes-Borrego M, et al. (2011) Molecular and pathogenic characterization of Fusarium redolens, a new causal agent of fusarium yellows in chickpea. Plant Dis 95: 860-870. https://doi.org/10.1094/PDIS-12-10-0946
|
| [5] |
Landa BB, Navas-Cortés JA, Jiménez-Díaz RM (2004) Integrated management of fusarium wilt of chickpea with sowing date, host resistance, and biological control. Phytopathology 94: 946-960. https://doi.org/10.1094/PHYTO.2004.94.9.946
|
| [6] |
Jiménez-Díaz RM, Castillo P, del Mar Jiménez-Gasco M, et al. (2015) Fusarium wilt of chickpeas: Biology, ecology and management. Crop Prot 73: 16-27. https://doi.org/10.1016/j.cropro.2015.02.023
|
| [7] |
Wise KA, Bradley CA, Pasche JS, et al. (2009) Resistance to QoI fungicides in Ascochyta rabiei from chickpea in the Northern Great Plains. Plant Dis 93: 528-536. https://doi.org/10.1094/PDIS-93-5-0528
|
| [8] |
Kouki H, Souihi M, Saadouli I, et al. (2003) Biocontrol potential of some rhizospheric soil bacterial strains against Fusarium culmorum and subsequent effect on growth of two tunisian wheat cultivars. Microorganisms 11: 1165-1165. https://doi.org/10.3390/microorganisms11051165
|
| [9] |
Hemissi I, Mabrouk Y, Mejri S, et al. (2013) Enhanced defence responses of chickpea plants against rhizoctonia solani by pre-inoculation with rhizobia. J Phytopathol 161: 412-418. https://doi.org/10.1111/jph.12071
|
| [10] |
Khan N, Martínez-Hidalgo P, Ice TA, et al. (2018) Antifungal activity of bacillus species against fusarium and analysis of the potential mechanisms used in biocontrol. Front Microbiol 9: 2363-2363. https://doi.org/10.3389/fmicb.2018.02363
|
| [11] |
Alekhya G, Gopalakrishnan S (2017) Biological control and plant growth-promotion traits of Streptomyces species under greenhouse and field conditions in chickpea. Agricul Res 6: 410-420. https://doi.org/10.1007/s40003-017-0278-2
|
| [12] | Khakipour N, Khavazi K, Mojallali H, et al. (2008) Production of auxin hormone by fluorescent pseudomonads. Am Eurasian J Agric Environ Sci 4: 687-692. http://www.idosi.org/aejaes/jaes4(6)/5.pdf |
| [13] |
Kumari S, Khanna V (2020) Induction of systemic resistance in chickpea (Cicer arietinum L.) against Fusarium oxysporum f. sp. ciceris by antagonistic rhizobacteria in assistance with native mesorhizobium. Curr Microbiol 77: 85-98. https://doi.org/10.1007/s00284-019-01805-6
|
| [14] |
Cachinero JM, Hervas A, Jiménez-Díaz RM, et al. (2002) Plant defence reactions against fusarium wilt in chickpea induced by incompatible race 0 of Fusarium oxysporum f. sp. ciceris and nonhost isolates of F. oxysporum. Plant Pathol 51: 765-776. https://doi.org/10.1046/j.1365-3059.2002.00760.x
|
| [15] |
Travers RS, Martin PAW, Reichelderfer CF (1987) Selective process for efficient isolation of soil Bacillus sp. Appl Environ Microbiol 53: 1263-1266. https://doi.org/10.1128/aem.53.6.1263-1266.1987
|
| [16] |
Kasana RC, Salwan R, Dhar H, et al. (2008) A rapid and easy method for the detection of microbial cellulases on agar plates using Gram's iodine. Curr Microbiol 57: 503-507. https://doi.org/10.1007/s00284-008-9276-8
|
| [17] |
Abdelwahed S, Saadouli I, Kouidhi S, et al. (2022) A new pioneer colorimetric micro-plate method for the estimation of ammonia production by plant growth promoting rhizobacteria (PGPR). Main Group Chem 21: 55-68. https://doi.org/10.3233/MGC-210077
|
| [18] |
Bric JM, Bostock RM, Silverstone SE (1991) Rapid in situ assay for indoleacetic Acid production by bacteria immobilized on a nitrocellulose membrane. Appl Environ Microbiol 57: 535-538.
|
| [19] |
Valérie Gravel, Hani Antoun, Russell J (2007) Tweddell, Growth stimulation and fruit yield improvement of greenhouse tomato plants by inoculation with Pseudomonas putida or Trichoderma atroviride: Possible role of indole acetic acid (IAA). Soil Biol Biochem 39: 1968-1977. https://doi.org/10.1016/j.soilbio.2007.02.015
|
| [20] |
Nautiyal CS (1999) An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol Lett 170: 265-270.
|
| [21] |
Fiske CH, Subbarow Y (1925) The colorimetric determination of phosphorus. J Biol Chem 66: 375-400. https://doi.org/10.1016/S0021-9258(18)84756-1
|
| [22] |
Grobelak A, Napora A, Kacprzak M (2015) Using plant growth-promoting rhizobacteria (PGPR) to improve plant growth. Ecol Engi 84: 22-28. https://doi.org/10.1016/j.ecoleng.2015.07.019
|
| [23] |
Payne SM (1994) Detection, isolation, and characterization of siderophores. Methods Enzymol 235: 329-44.
|
| [24] |
Olsen S, Sommers L (1982) Phosphorus. Methods of soil analysis: Part 2. chemical and microbiological properties . the American Society of Agronomy, Inc., Soil Science Society of America, Inc 403-430. https://doi.org/10.2134/agronmonogr9.2.2ed
|
| [25] |
Palmieri D, Vitullo D, De Curtis F, et al. (2017) A microbial consortium in the rhizosphere as a new biocontrol approach against Fusarium decline of chickpea. Plant Soil 412: 425-439. https://doi.org/10.1007/s11104-016-3080-1
|
| [26] |
Vejan P, Abdullah R, Khadiran T, et al. (2016) Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules 21: 573-573. https://doi.org/10.3390/molecules21050573
|
| [27] |
Kaur R, Singh RS, Alabouvette C (2007) Antagonistic activity of selected isolates of fluorescent Pseudomonas against Fusarium oxysporum f.sp. ciceris. Asian J Pl Sci 6: 446-456. http://www.ansinet.org/ajps
|
| [28] |
Arfaoui A, El Hadrami A, Mabrouk Y, et al. (2007) Treatment of chickpea with Rhizobium isolates enhances the expression of phenylpropanoid defense-related genes in response to infection by Fusarium oxysporum f. sp. ciceris. Plant Physiol Biochem 45: 470-479. https://doi.org/10.1016/j.plaphy.2007.04.004
|
| [29] |
Khalifa M, Rouag N, Bouhadida M (2022) Evaluation of the antagonistic effect of pseudomonas rhizobacteria on Fusarium wilt of chickpea. Agriculture 12: 429-429. https://doi.org/10.3390/agriculture12030429
|
| [30] | Jadhav HP, Shaikh SS, Sayyed RZ (2017) Role of hydrolytic enzymes of rhizoflora in biocontrol of fungal phytopathogens: An Overview. Rhizotrophs: Plant Growth Promotion to Bioremediation. Microorganisms for Sustainability . Singapore: Springer. https://doi.org/10.1007/978-981-10-4862-3_9 |
| [31] |
Sindhu S, Research KDM (2001) Chitinolytic and cellulolytic Pseudomonas sp. antagonistic to fungal pathogens enhances nodulation by Mesorhizobium sp. Cicer in chickpea. Microbiol Res 156: 353-358. https://doi.org/10.1078/0944-5013-00120
|
| [32] |
Vlassi A, Nesler A, Perazzolli M, et al. (2020) Volatile organic compounds from Lysobacter capsici AZ78 as potential candidates for biological control of soilborne plant pathogens. Front Microbiol 11: 1748-1748. https://doi.org/10.3389/fmicb.2020.01748
|
| [33] |
Sreevidya M, Gopalakrishnan S, Melo TM, et al. (2015) Biological control of Botrytis Cinerea and plant growth promotion potential by Penicillium Citrinum in chickpea (Cicer arietinum L). Biocontrol Sci Technol Taylor Francis 25: 739-55. https://doi.org/10.1080/09583157.2015.1010483
|
| [34] |
Dobbelaere SJ, Vanderleyden Okon Y (2003) Plant growth promoting effects of diazotrophs in the rhizosphere. Crit Rev Plant Sci 22: 107-149. https://doi.org/10.1080/713610853
|
| [35] |
Castagno LN, Estrella MJ, Sannazzaro AI, et al. (2011) Phosphate solubilization mechanism and in vitro plant growth promotion activity mediated by Pantoea eucalypti isolated from Lotus tenuis rhizosphere in the Salado River Basin (Argentina). J Appl Microbiol 110: 1151-1165. https://doi.org/10.1111/j.1365-2672.2011.04968.x
|
| [36] |
Valverde A, Burgos A, Fiscella T, et al. (2006) Differential effects of coinoculations with Pseudomonas jessenii PS06 (a phosphate-solubilizing bacterium) and Mesorhizobium ciceri C-2/2 strains on the growth and seed yield of chickpea under greenhouse and field conditions. Plant Soil 287: 43-50. https://doi.org/10.1007/s11104-006-9057-8
|
| [37] |
Rudresh DL, Shivaprakash MK, Prasad RD (2005) Effect of combined application of Rhizobium, phosphate-solubilizing bacterium, and Trichoderma spp. on growth, nutrient uptake, and yield of chickpea (Cicer aritenium L.). Appl Soil Ecol 28: 139-146. https://doi.org/10.1016/j.apsoil.2004.07.005
|
| [38] |
Sindhu SS, Suneja S, Goel AK, et al. (2002) Plant growth–promoting effects of Pseudomonas sp. on coinoculation with Mesorhizobium sp. cicer strain under sterile and “wilt sick” soil conditions. Appl Soil Ecol 19: 57-64. https://doi.org/10.1016/S0929-1393(01)00176-7
|
| [39] |
Alagawadi AR, Gaur AC (1988) Associative effect of Rhizobium and phosphate-solubilizing bacteria on the yield and nutrient uptake of chickpea. Plant Soil 105: 241-246. https://doi.org/10.1007/BF02376788
|
| [40] | Kumar R, Chandra R (2008) Influence of PGPR and PSB on Rhizobium leguminosarum bv. viciae strain competition and symbiotic performance in lentil. World J Agric Sci 4: 297-301. http://www.idosi.org/wjas/wjas4(3)/2.pdf |
| [41] |
Tilak KVBR, Ranganayaki N, Manoharachari C (2006) Synergistic effects of plant growth– promoting rhizobacteria and Rhizobium on nodulation and nitrogen fixation by pigeonpea (Cajanus cajan). Eur J Soil Sci 57: 67-71. https://doi.org/10.1111/j.1365-2389.2006.00771.x
|
| [42] |
Zaidi A, Khan MS, Amil M (2003) Interactive effect of rhizotrophic microorganisms on yield and nutrient uptake of chickpea (Cicer arietinum L.). Eur J Agronomy 19: 15-21. https://doi.org/10.1016/S1161-0301(02)00015-1
|
| [43] |
Verma JP, Yadav J, Tiwari KN (2012) Enhancement of nodulation and yield of chickpea by co-inoculation of indigenous Mesorhizobium spp. and plant growth-promoting rhizobacteriain eastern Uttar Pradesh. Commun. Soil Sci Plant Anal 43: 605-621.
|
| [44] |
Barbosa JM, Hungria M, Prior SA, et al. (2022) Improving yield and health of legume crops via co-inoculation with rhizobia and Trichoderma: A global meta-analysis. Appl Soil Ecol 176: 104493.
|
| [45] | Zian AH, Aly MM (2020) Impact of Co-Inoculation with Rhizobium leguminosarum and some Plant Growth Promoting Rihzobacteria against Rhizoctonia solani and Fusarium oxysporum Infected Faba Bean. J Plant Prot Pathol 11: 441-453. |
| [46] |
Gamalero E, Glick BR (2011) Mechanisms used by plant growth promoting bacteria. Bacteria in agrobiology: plant nutrient management . Berlin, Heidelberg: Springer 17-46.
|
| [47] | Bacon CW, Hinton DM (2002) Endophytic and biological control potential of Bacillus mojavensis and related species. BiolControl 23: 274-84. |
| [48] |
Ossowicki A, Jafra S, Garbeva P (2017) The antimicrobial volatile power of the rhizospheric isolate Pseudomonas donghuensis P482. PLOS One 12: e0174362. https://doi.org/10.1371/journal.pone.0174362
|
| [49] |
Munjal V, Nadakkakath AV, Sheoran N, et al. (2016) Genotyping and identification of broad-spectrum antimicrobial volatiles in black pepper root endophytic biocontrol agent, Bacillus megaterium BP17: theory and applications in pest management. Biol Control 92: 66-76.
|
| [50] |
Nithyapriya S, Sundaram L, Eswaran S, et al. (2024) Purification and characterization of desferrioxamine b of Pseudomonas fluorescens and its application to improve oil content, nutrient uptake, and plant growth in peanuts. Microb Ecol 87: 60. https://doi.org/10.1007/s00248-024-02377-0
|
| [51] |
Sharma S, Kumari P, Shandilya M, et al. (2025) The combination of α-Fe2O3 NP and Trichoderma sp. improves antifungal activity against fusarium wilt. J Basic Microbiol 19: e2400613. https://doi.org/10.1002/jobm.202400613
|

Figures(4) / Tables(5)

Sabrine Balti, Yassine Mabrouk, Mouna Souihi, Imen Hemissi, Ismail Amri, Ethan Humm, Noor Khan, Ann M. Hirsch. Combined inoculation of rhizobacteria with Mesorhizobium promotes growth, nutrient contents, and protects chickpea against Fusarium redolens[J]. AIMS Microbiology, 2025, 11(2): 318-337. doi: 10.3934/microbiol.2025015







 DownLoad:
DownLoad: