Drug repositioning has emerged as a promising strategy for assessing its antimicrobial efficacy in treating infectious diseases.
Seventy-five samples were collected and investigated for the presence of Pseudomonas aeruginosa. Antibiotic resistance, hemolytic activity, twitching motility, and biofilm formation were assessed. lasI and lasR genes were detected using conventional PCR. Minimum inhibitory concentrations of paroxetine, fluoxetine, and levofloxacin were determined by broth micro-dilution. The fractional inhibitory concentration index was calculated to assess the interaction between fluoxetine/levofloxacin and paroxetine/levofloxacin combinations. Half the MIC values of the drugs were selected for inhibitory effect assessment for virulence factors. Antibacterial and healing effects of fluoxetine were investigated on 30 male albino rats using a digital camera, bacterial count, and histological examination.
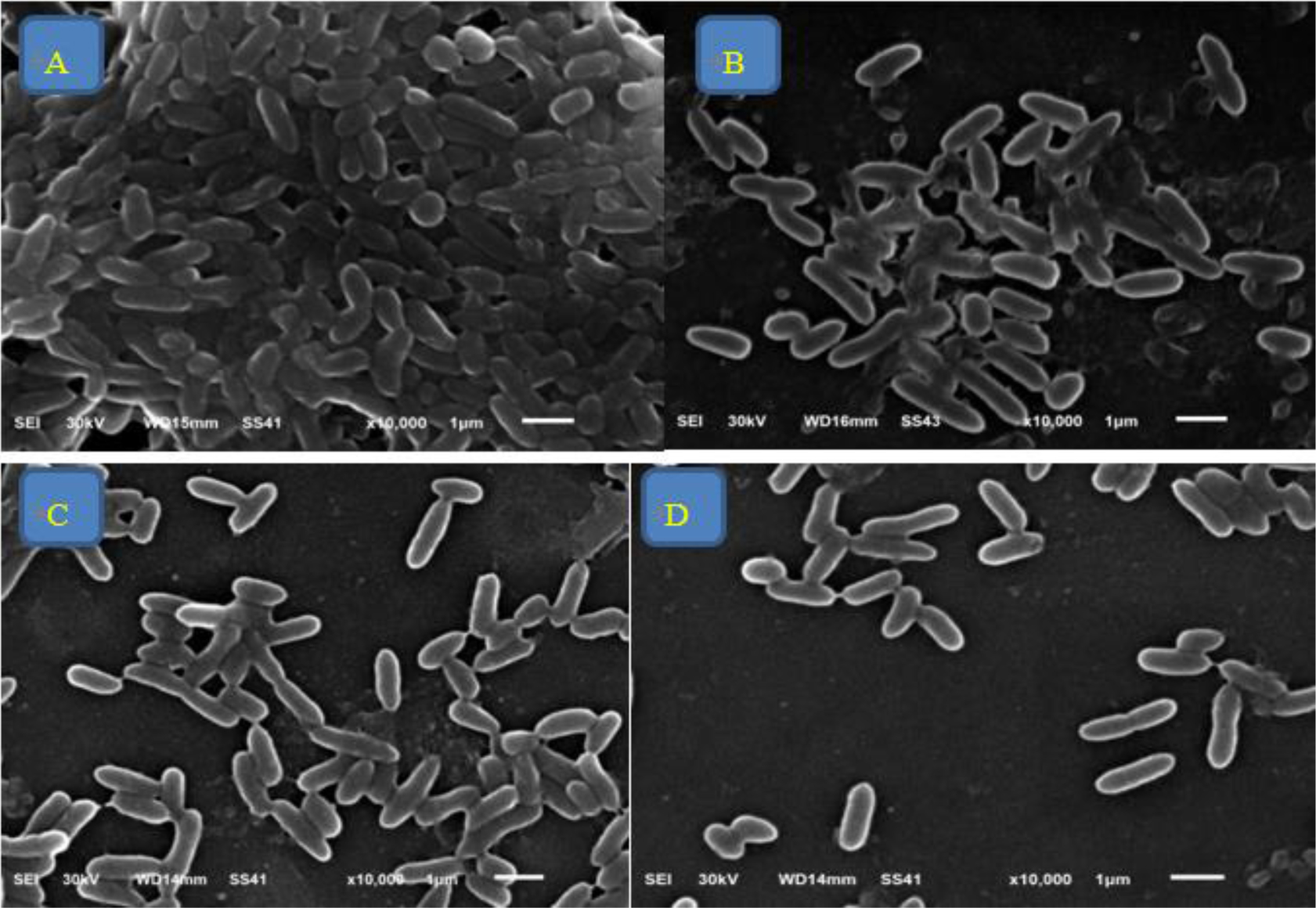
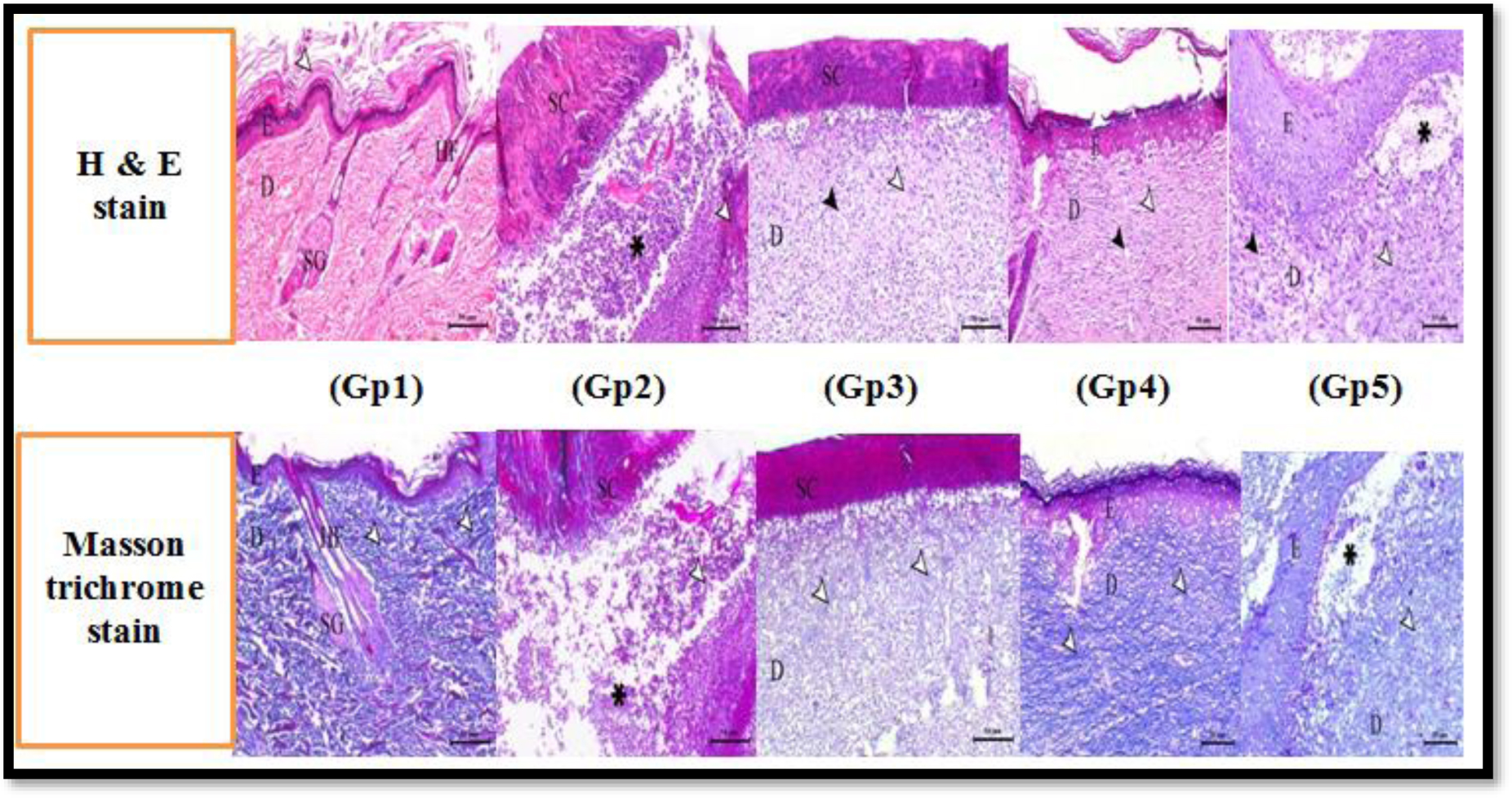
Our 25 P. aeruginosa isolates were highly drug-resistant. 80%, 92%, and 80% of isolates were positive for twitching motility, hemolysis, and biofilm formation, respectively. 92% of isolates were positive for lasI gene and 96% for lasR gene. MICs of fluoxetine and paroxetine ranged from 32 to 512 µg/mL and MICs of levofloxacin ranged from 1 to 256 µg/mL. A synergistic outcome was observed in both combinations. Biofilm formation, twitching motility, and hemolysis were inhibited by paroxetine and fluoxetine in the majority of isolates. Fluoxetine/levofloxacin and paroxetine/levofloxacin combinations inhibited twitching motility, hemolysis, and biofilm formation in all isolates. Enhanced wound healing was observed in rats treated with fluoxetine and levofloxacin, with the fluoxetine/levofloxacin combination group demonstrating the most significant wound-healing effect. Bacterial count decreased in rats treated with levofloxacin, fluoxetine, and the levofloxacin/fluoxetine combination. Histological examination revealed higher wound healing in the levofloxacin-treated group than the fluoxetine group, and the combination treatment group displayed the fastest rate of wound healing.
Paroxetine and fluoxetine showed considerable antibacterial inhibitory effects against multi-drug resistant P. aeruginosa isolates. Fluoxetine showed significant improvement in anti-inflammatory effects and wound healing. To the best of our knowledge, this is the first Egyptian study to investigate the repurposing of paroxetine and fluoxetine as antibacterial agents. Further studies are needed to investigate their applicability as antibacterial agents as single agents or in combination with other antibiotics.
Citation: Kholoud Baraka, Rania Abozahra, Eman Khalaf, Mahmoud Elsayed Bennaya, Sarah M. Abdelhamid. Repurposing of paroxetine and fluoxetine for their antibacterial effects against clinical Pseudomonas aeruginosa isolates in Egypt[J]. AIMS Microbiology, 2025, 11(1): 126-149. doi: 10.3934/microbiol.2025007
Drug repositioning has emerged as a promising strategy for assessing its antimicrobial efficacy in treating infectious diseases.
Seventy-five samples were collected and investigated for the presence of Pseudomonas aeruginosa. Antibiotic resistance, hemolytic activity, twitching motility, and biofilm formation were assessed. lasI and lasR genes were detected using conventional PCR. Minimum inhibitory concentrations of paroxetine, fluoxetine, and levofloxacin were determined by broth micro-dilution. The fractional inhibitory concentration index was calculated to assess the interaction between fluoxetine/levofloxacin and paroxetine/levofloxacin combinations. Half the MIC values of the drugs were selected for inhibitory effect assessment for virulence factors. Antibacterial and healing effects of fluoxetine were investigated on 30 male albino rats using a digital camera, bacterial count, and histological examination.
Our 25 P. aeruginosa isolates were highly drug-resistant. 80%, 92%, and 80% of isolates were positive for twitching motility, hemolysis, and biofilm formation, respectively. 92% of isolates were positive for lasI gene and 96% for lasR gene. MICs of fluoxetine and paroxetine ranged from 32 to 512 µg/mL and MICs of levofloxacin ranged from 1 to 256 µg/mL. A synergistic outcome was observed in both combinations. Biofilm formation, twitching motility, and hemolysis were inhibited by paroxetine and fluoxetine in the majority of isolates. Fluoxetine/levofloxacin and paroxetine/levofloxacin combinations inhibited twitching motility, hemolysis, and biofilm formation in all isolates. Enhanced wound healing was observed in rats treated with fluoxetine and levofloxacin, with the fluoxetine/levofloxacin combination group demonstrating the most significant wound-healing effect. Bacterial count decreased in rats treated with levofloxacin, fluoxetine, and the levofloxacin/fluoxetine combination. Histological examination revealed higher wound healing in the levofloxacin-treated group than the fluoxetine group, and the combination treatment group displayed the fastest rate of wound healing.
Paroxetine and fluoxetine showed considerable antibacterial inhibitory effects against multi-drug resistant P. aeruginosa isolates. Fluoxetine showed significant improvement in anti-inflammatory effects and wound healing. To the best of our knowledge, this is the first Egyptian study to investigate the repurposing of paroxetine and fluoxetine as antibacterial agents. Further studies are needed to investigate their applicability as antibacterial agents as single agents or in combination with other antibiotics.

| [1] |
Roca I, Akova M, Baquero F, et al. (2015) The global threat of antimicrobial resistance: science for intervention. New Microbes New Infect 6: 22-29. https://doi.org/10.1016/j.nmni.2015.02.007 
|
| [2] |
Spellberg B, Srinivasan A, Chambers HF (2016) New societal approaches to empowering antibiotic stewardship. JAMA 315: 1229-1230. https://doi.org/10.1001/jama.2016.1346
|
| [3] | (2015) World Health OrganizationGlobal action plan on antimicrobial resistance. Geneva: World Health Organization. https://www.who.int/antimicrobial-resistance/global-action-plan/en/ |
| [4] | Centers for Disease Control and PreventionAntibiotic use in the United States: progress and opportunities, Atlanta (2017). https://www.cdc.gov/antibiotic-use/stewardship-report/pdf/stewardship-report.pdf |
| [5] |
Wellington EMH, Boxall ABA, Cross P (2013) The role of the natural environment in the emergence of antibiotic resistance in Gram-negative bacteria. Lancet Infect Dis 13: 155-165. https://doi.org/10.1016/S1473-3099(12)70317-1
|
| [6] |
Morehead MS, Scarbrough C (2018) Emergence of global antibiotic resistance. Prim Care 45: 467-484. https://doi.org/10.1016/j.pop.2018.05.006
|
| [7] |
Brown D (2015) Antibiotic resistance breakers: can repurposed drugs fill the antibiotic discovery void?. Nat Rev Drug Discov 14: 821-832. https://doi.org/10.1038/nrd4675
|
| [8] |
Yang Y, Zhang Z, Li S, et al. (2014) Synergy effects of herb extracts: pharmacokinetics and pharmacodynamic basis. Fitoterapia 92: 133-147. https://doi.org/10.1016/j.fitote.2013.10.010
|
| [9] | Kaur I (2016) Novel strategies to combat antimicrobial resistance. J Infect Dis Ther 4: 4. https://doi.org/10.4172/2332-0877.1000292 |
| [10] |
Serafin MB, Hörner R (2018) Drug repositioning, a new alternative in infectious diseases. Braz J Infect Dis 22: 252-256. https://doi.org/10.1016/j.bjid.2018.05.007
|
| [11] |
Ashburn TT, Thor KB (2004) Drug repositioning: identifying and developing new uses for existing drugs. Nat Rev Drug Discov 3: 673-683. https://doi.org/10.1038/nrd1468
|
| [12] |
Pushpakom S, Iorio F, Eyers PA, et al. (2018) Drug repurposing: progress, challenges and recommendations. Nat Rev Drug Discov 18: 41-58. https://doi.org/10.1038/nrd.2018.168
|
| [13] |
Xue H, Li J, Xie H, et al. (2018) Review of drug repositioning approaches and resources. Int J Biol Sci 14: 1232-1244. https://doi.org/10.7150/ijbs.24612
|
| [14] |
Gelosa P, Castiglioni L, Camera M, et al. (2020) Drug repurposing in cardiovascular diseases: opportunity or hopeless dream?. Biochem Pharmacol 177: 113894. https://doi.org/10.1016/j.bcp.2020.113894
|
| [15] |
Rastelli G, Pellati F, Pinzi L, et al. (2020) Repositioning natural products in drug discovery. Molecules 25: 1154. https://doi.org/10.3390/molecules25051154
|
| [16] |
Vogt I, Mestres J (2010) Drug-Target Networks. Mol Inform 29: 10-14. https://doi.org/10.1002/minf.200900069
|
| [17] |
De Sousa AK, Rocha JE, de Souza TG, et al. (2018) New roles of fluoxetine in pharmacology: Antibacterial effect and modulation of antibiotic activity. Microb Pathog 123: 368-371. https://doi.org/10.1016/j.micpath.2018.07.040
|
| [18] |
Munoz-Bellido JL, Munoz-Criado S, Garca-Rodrguez J (2000) Antimicrobial activity of psychotropic drugs selective serotonin reuptake inhibitors. Int J Antimicrob Agents 14: 177-180. https://doi.org/10.1016/S0924-8579(99)00154-5
|
| [19] |
Foletto VS, Serafin MB, da Rosa TF, et al. (2019) Fluoxetine and paroxetine: repositioning as a therapeutic alternative in the treatment of various diseases. Am J Ther 27: 547-551. https://doi.org/10.1097/MJT.0000000000001011
|
| [20] |
Gosain A, DiPietro L (2004) Aging and wound healing. World J Surg 28: 321-326. https://doi.org/10.1007/s00268-003-7397-6
|
| [21] |
Mathieu D, Linke J, Wattel F (2006) Non-healing wounds. Handbook on hyperbaric medicine . New York: Springer 401-27. https://doi.org/10.1007/1-4020-4448-8_20
|
| [22] |
Menke N, Ward K, Witten T, et al. (2007) Impaired wound healing. Clin Dermatol 25: 19-25. https://doi.org/10.1016/j.clindermatol.2006.12.005
|
| [23] |
Malinin A, Oshrine B, Serebruany V (2004) Treatment with selective serotonin reuptake inhibitors for enhancing wound healing. Med Hypothese 63: 103-109. https://doi.org/10.1016/j.mehy.2003.10.021
|
| [24] | Bailey WR, Scott EG (1986) Bailey and Scott's Diagnostic Microbiology. Mosby. Chapter 7 |
| [25] |
Bauer AW, Kirby WM, Sherris JC (1966) Antibiotic susceptibility testing by a standardized single disk method. Am J Clin Pathol 45: 493-496. https://doi.org/10.1093/ajcp/45.4_ts.493
|
| [26] | Clinical and Laboratory Performance Standards For Antimicrobial Susceptibility Testing - 30th Edition, 2020. https://webstore.ansi.org/standards/clsi/clsim100ed30 |
| [27] |
Castro-Escarpulli G, Figueras MJ, Aguilera-Arreola G, et al. (2003) Characterisation of Aeromonas spp. isolated from frozen fish intended for human consumption in Mexico. Int J Food Microbiol 84: 41-49. https://doi.org/10.1016/S0168-1605(02)00393-8
|
| [28] |
Kus JV, Tullis E, Cvitkovitch DG, et al. (2004) Significant differences in type IV pilin allele distribution among Pseudomonas aeruginosa isolates from cystic fibrosis (CF) versus non-CF patients. Microbiology 150: 1315-1326. https://doi.org/10.1099/mic.0.26822-0
|
| [29] |
Freeman J, Falkiner FR, Keane CT (1989) New method for detecting slime production by coagulase negative staphylococci. J Clin Pathol 42: 872-874. https://doi.org/10.1136/jcp.42.8.872
|
| [30] | O'Toole GAJJ (2011) Microtiter dish biofilm formation assay. J Vis Exp 47: e2437. https://doi.org/10.3791/2437 |
| [31] |
Queipo-Ortun MI, Colmenero JDD, Macias M, et al. (2008) Preparation of bacterial DNA template by boiling and effect of immunoglobulin G as an inhibitor in real-time PCR for serum samples from patients with brucellosis. Clin Vaccine Immunol 15: 293-296. https://doi.org/10.1128/CVI.00270-07
|
| [32] |
Karatuna O, Yagci A (2010) Analysis of quorum sensing-dependent virulence factor production and its relationship with antimicrobial susceptibility in Pseudomonas aeruginosa respiratory isolates. Clin Microbiol Infect 16: 1770-1775. https://doi.org/10.1111/j.1469-0691.2010.03177.x
|
| [33] |
Wiegand I, Hilpert K, Hancock WER (2008) Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat Protoc 3: 163-175. https://doi.org/10.1038/nprot.2007.521
|
| [34] |
Konaté K, Movoungou JF, Lepengué AN, et al. (2012) Antibacterial activity against β-lactamase producing Methicillin and Ampicillin-resistant Staphylococcus aureus: fractional Inhibitory Concentration Index (FICI) determination. Ann Clin Microbiol Antimicrob 11: 18. https://doi.org/10.1186/1476-0711-11-18
|
| [35] |
Odds FC (2003) Synergy, antagonism, and what the chequerboard puts between them. J Antimicrob Chemother 52: 1. https://doi.org/10.1093/jac/dkg301
|
| [36] | Fassel TA, Edmiston CE (2000) Evaluating adherent bacteria and biofilm using electron microscopy. Handbook of Bacterial Adhesion: Principles, Methods, and Applications . New Jersey: Humana Press Inc 235-248. https://doi.org/10.1007/978-1-59259-224-1_14 |
| [37] |
Yu N, Wang X, Qiu L, et al. (2020) Bacteria-triggered hyaluronan/AgNPs/gentamicin nanocarrier for synergistic bacteria disinfection and wound healing application. Chem Eng J 380: 122582. https://doi.org/10.1016/j.cej.2019.122582
|
| [38] |
Shu G, Xu D, Zhang W, et al. (2022) Preparation of shikonin liposome and evaluation of its in-vitro antibacterial and in-vivo infected wound healing activity. Phytomedicine 99: 154035. https://doi.org/10.1016/j.phymed.2022.154035
|
| [39] |
Magiorakos AP, Srinivasan A, Carey RB, et al. (2012) Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect 18: 268-281. https://doi.org/10.1111/j.1469-0691.2011.03570.x
|
| [40] |
Gaynes R, Edwards JR (2005) Overview of nosocomial infections caused by gram-negative bacilli. Clin Infect Dis 41: 848-54. https://doi.org/10.1086/432803
|
| [41] | Diab M, Fam N, El-Said M, et al. (2013) Occurrence of VIM-2 metallo-β-lactamases in imipenem resistant and susceptible Pseudomonas aeruginosa clinical isolates from Egypt. Afr J Microbiol Res 7: 4465-4472. https://doi.org/10.5897/AJMR2013.6181 |
| [42] |
Abbas HA, El-Ganiny AM, Kamel HA (2018) Phenotypic and genotypic detection of antibiotic resistance of Pseudomonas aeruginosa isolated from urinary tract infections. Afr Health Sci 18: 11-21. https://doi.org/10.4314/ahs.v18i1.3
|
| [43] |
Rodulfo H, Arcia A, Hernandez A, et al. (2019) Virulence factors and integrons are associated with MDR and XDR phenotypes in nosocomial strains of Pseudomonas aeruginosa in a Venezuelan university hospital. Rev Inst Med Trop Sao Paulo 61: e20. https://doi.org/10.1590/s1678-9946201961020
|
| [44] |
Rossi Goncalves I, Dantas RCC, Ferreira ML, et al. (2017) Carbapenem-resistant Pseudomonas aeruginosa: association with virulence genes and biofilm formation. Braz J Microbiol 48: 211-217. https://doi.org/10.1016/j.bjm.2016.11.004
|
| [45] |
Hashem H, Hanora A, Abdalla S, et al. (2017) Dissemination of metallo-beta-lactamase in Pseudomonas aeruginosa isolates in Egypt: mutation in blaVIM-4. APMIS 125: 0499-0505. https://doi.org/10.1111/apm.12669
|
| [46] |
Sonbol FI, Khalil MAEF, Mohamed AB, et al. (2015) Correlation between antibiotic resistance and virulence of Pseudomonas aeruginosa clinical isolates. Turk J Med Sci 45: 568-577. https://doi.org/10.3906/sag-1406-58
|
| [47] |
Zhu H, Bandara R, Conibear TC, et al. (2004) Pseudomonas aeruginosa with lasI quorum-sensing deficiency during corneal infection. Invest Ophthalmol Vis Sci 45: 1897-1903. https://doi.org/10.1167/iovs.03-0980
|
| [48] |
Dogonchi AA, Ghaemi EA, Ardebili A, et al. (2018) Metallo-beta-lactamase-mediated resistance among clinical carbapenem-resistant Pseudomonas aeruginosa isolates in northern Iran: a potential threat to clinical therapeutics. Tzu Chi Med J 30: 90-96. https://doi.org/10.4103/tcmj.tcmj_101_17
|
| [49] |
Al Dawodeyah HY, Obeidat N, Abu-Qatouseh LF, et al. (2018) Antimicrobial resistance and putative virulence genes of Pseudomonas aeruginosa isolates from patients with respiratory tract infection. Germs 8: 31-40. https://doi.org/10.18683/germs.2018.1130
|
| [50] |
Baniya B, Pant ND, Neupane S, et al. (2017) Biofilm and metallo betalactamase production among the strains of Pseudomonas aeruginosa and Acinetobacter spp. at a Tertiary Care Hospital in Kathmandu, Nepal. Ann Clin Microbiol Antimicrob 16: 70. https://doi.org/10.1186/s12941-017-0245-6
|
| [51] |
Bogiel T, Deptuła A, Kwiecińska-piróg J, et al. (2017) The prevalence of exoenzyme S gene in multidrug-sensitive and multidrug-resistant Pseudomonas aeruginosa clinical strains. Pol J Microbiol 66: 427-431. https://doi.org/10.5604/01.3001.0010.6500
|
| [52] |
Ellappan K, Belgode Narasimha H, et al. (2018) Coexistence of multidrug resistance mechanisms and virulence genes in carbapenem-resistant Pseudomonas aeruginosa strains from a tertiary care hospital in South India. J Glob Antimicrob Resist 12: 37-43. https://doi.org/10.1016/j.jgar.2017.08.018
|
| [53] |
Ghanbarzadeh Corehtash Z, Khorshidi A, Firoozeh F, et al. (2015) Biofilm formation and virulence factors among Pseudomonas aeruginosa isolated from burn patients. Jundishapur J Microbiol 8: e22345. https://doi.org/10.5812/jjm
|
| [54] |
Wang H, Tu F, Gui Z, et al. (2013) Antibiotic Resistance Profiles and Quorum Sensing-Dependent Virulence Factors in Clinical Isolates of Pseudomonas aeruginosa. Indian J Microbiol 53: 163-167. https://doi.org/10.1007/s12088-013-0370-7
|
| [55] |
Deligianni E, Pattison S, Berrar D, et al. (2010) Pseudomonas aeruginosa cystic fibrosis isolates of similar RAPD genotype exhibit diversity in biofilm forming ability in vitro. BMC Microbiol 10: 38. https://doi.org/10.1186/1471-2180-10-38
|
| [56] | Ochoa S, López-Montiel F, Escalona G, et al. (2013) Pathogenic characteristics of Pseudomonas aeruginosa strains resistant to carbapenems associated with biofilm formation. Bol Med Hosp Infant Mex 70: 133-144. |
| [57] |
Aqel H, Sannan N, Foudah R, et al. (2023) Enzyme production and inhibitory potential of Pseudomonas aeruginosa: contrasting clinical and environmental isolates. Antibiotics (Basel) 12: 1354. https://doi.org/10.3390/antibiotics12091354
|
| [58] |
Senturk S, Ulusoy S, Bosgelmez-Tinaz G, et al. (2012) Quorum sensing and virulence of Pseudomonas aeruginosa during urinary tract infections. J Infect Dev Ctries 6: 501-507. https://doi.org/10.3855/jidc.2543
|
| [59] |
Edward EA, El Shehawy MR, Abouelfetouh A, et al. (2023) Prevalence of different virulence factors and their association with antimicrobial resistance among Pseudomonas aeruginosa clinical isolates from Egypt. BMC Microbiol 23: 161. https://doi.org/10.1186/s12866-023-02897-8
|
| [60] | Abd El Galil K, AbdelGhani S, Sebak M, et al. (2013) Detection of biofilm genes among clinical isolates of Pseudomonas aeruginosa recovered from some Egyptian hospitals. Egypt J Microbiol 36. |
| [61] |
El-Mahdy R, El-Kannishy G (2019) Virulence factors of Carbapenem-resistant Pseudomonas aeruginosa in hospital-acquired infections in Mansoura, Egypt. Infect Drug Resist 12: 3455-3461. https://doi.org/10.2147/IDR.S222329
|
| [62] |
Deptula A, Gospodarek E (2010) Reduced expression of virulence factors in multidrug-resistant Pseudomonas aeruginosa strains. Arch Microbiol 192: 79-84. https://doi.org/10.1007/s00203-009-0528-1
|
| [63] |
Ghanem SM, Abd El-Baky RM, Abourehab MAS, et al. (2023) Prevalence of quorum sensing and virulence factor genes among Pseudomonas aeruginosa isolated from patients suffering from different infections and their association with antimicrobial resistance. Infect Drug Resist 16: 2371-2385. https://doi.org/10.2147/IDR.S403441
|
| [64] | Sabharwal N, Dhall S, Chhibber S, et al. (2014) Molecular detection of virulence genes as markers in Pseudomonas aeruginosa isolated from urinary tract infections. Int J Mol Epidemiol Genet 5: 125-134. |
| [65] | Kadhim D, Ali Mi R (2014) Prevalence study of quorum sensing groups among clinical isolates of Pseudomonas aeruginosa. Int J Curr Microbiol App Sci 3: 204-215. https://doi.org/10.2147/IDR.S403441 |
| [66] | Aghamollaei H, Azizi BK, Moosazadeh MM (2014) Rapid detection of Pseudomonas aeruginosa by PCR method using specific primers of quorum sensing LasI gene. Armaghane Danesh 18: 723-735. http://armaghanj.yums.ac.ir/article-1-480-en.html |
| [67] |
Sousa DKA, Rocha EJ, Souza DGT, et al. (2018) New roles of fluoxetine in pharmacology: Antibacterial effect and modulation of antibiotic activity. Microbial Pathogenesis 123: 368-371. https://doi.org/10.1016/j.micpath.2018.07.040
|
| [68] |
Foletto SV, da Rosa FT, Serafin BM, et al. (2021) Repositioning of antidepressant drugs and synergistic effect with ciprofloxacin against multidrug-resistant bacteria. World J Microbiol Biotechnol 37. https://doi.org/10.1007/s11274-021-03016-3
|
| [69] |
Guedes GM, Araújo ES, Ribeiro KV, et al. (2023) Effect of fluoxetine on planktonic and biofilm growth and the antimicrobial susceptibility of Burkholderia pseudomallei. Future Microbiol 18: 785-794. https://doi.org/10.2217/fmb-2022-0272
|
| [70] |
Pereira TC, de Menezes RT, de Oliveira HC, et al. (2021) In vitro synergistic effects of fluoxetine and paroxetine in combination with amphotericin B against Cryptococcus neoformans. Pathog Dis 79: ftab001. https://doi.org/10.1093/femspd/ftab001
|
| [71] |
Kiecka A, Szczepanik M (2022) The potential action of SSRIs in the treatment of skin diseases including atopic dermatitis and slow-healing wounds. Pharmacol Rep 74: 947-955. https://doi.org/10.1007/s43440-022-00423-7
|
| [72] |
Patel DK, Ganguly K, Hexiu J, et al. (2022) Functionalized chitosan/spherical nanocellulose-based hydrogel with superior antibacterial efficiency for wound healing. Carbohydr Polym 284: 119202. https://doi.org/10.1016/j.carbpol.2022.119202
|
| [73] |
Farahani RM, Sadr K, Rad JS, et al. (2007) Fluoxetine enhances cutaneous wound healing in chronically stressed Wistar rats. Adv Skin Wound Care 20: 157-165. https://doi.org/10.1097/01.ASW.0000262710.59293.6b
|
| [74] |
Yuksel EP, Ilkaya F, Yildiz L, et al. (2014) Effects of paroxetine on cutaneous wound healing in healthy and diabetic rats. Adv Skin Wound Care 27: 216-221. https://doi.org/10.1097/01.ASW.0000445920.14039.64
|
| [75] | Dwajani S, Gurumurthy R (2013) Wound healing property of paroxetine in immunosuppressed albino rats. IJBCP 2: 772. https://doi.org/10.5455/2319-2003.ijbcp20131219 |
| [76] |
Fazzino F, Montes C, Urbina M, et al. (2008) Serotonin transporter is differentially localized in subpopulations of lymphocytes of major depression patients. Effect of fluoxetine on proliferation. J Neuroimmunol 196: 173. https://doi.org/10.1016/j.jneuroim.2008.03.012
|
| [77] |
Hou R, Ye G, Liu Y, et al. (2019) Effects of SSRIs on peripheral inflammatory cytokines in patients with generalized anxiety disorder. Brain Behave Immun 81: 105-110. https://doi.org/10.1016/j.bbi.2019.06.001
|

Figures(3) / Tables(6)

Kholoud Baraka, Rania Abozahra, Eman Khalaf, Mahmoud Elsayed Bennaya, Sarah M. Abdelhamid. Repurposing of paroxetine and fluoxetine for their antibacterial effects against clinical Pseudomonas aeruginosa isolates in Egypt[J]. AIMS Microbiology, 2025, 11(1): 126-149. doi: 10.3934/microbiol.2025007







 DownLoad:
DownLoad: