Citation: Jenise M. Bauman, Sarah Francino, Amy Santas. Interactions between ectomycorrhizal fungi and chestnut blight (Cryphonectria parasitica) on American chestnut (Castanea dentata) used in coal mine restoration[J]. AIMS Microbiology, 2018, 4(1): 104-122. doi: 10.3934/microbiol.2018.1.104

| [1] | Smith SE, Read DJ (2008) Mycorrhizal Symbiosis, 3 Eds., San Diego: Academic Press. |
| [2] |
Bever JD, Dickie IA, Facelli E, et al. (2010) Rooting theories of plant community ecology in microbial interactions. Trends Ecol Evol 25: 468–478. doi: 10.1016/j.tree.2010.05.004

|
| [3] |
Zipper CE, Burger JA, Skousen JG, et al. (2011) Restoring forests and associated ecosystem services on Appalachian coal surface mines. Environ Manag 47: 751–765. doi: 10.1007/s00267-011-9670-z
|
| [4] |
Simard SW, Durall DM (2004) Mycorrhizal networks: A review of their extent, function and importance. Can J Bot 82: 1140–1165. doi: 10.1139/b04-116
|
| [5] | Van Der Heijden MGA, Horton TR (2009) Socialism in soil? The importance of mycorrhizal fungal networks for facilitation in natural ecosystems. J Ecol 97: 1139–1150. |
| [6] |
Mangan SA, Schnitzer SA, Herre EA, et al. (2010) Negative plant-soil feedback predicts tree-species relative abundance in a tropical forest. Nature 466: 752–755. doi: 10.1038/nature09273
|
| [7] |
Bever JD, Mangan SA, Alexander HM (2015) Maintenance of plant species diversity by pathogens. Annu Rev Ecol Evol S 46: 305–325. doi: 10.1146/annurev-ecolsys-112414-054306
|
| [8] |
Marloes H, Liesje M, Hannie C, et al. (2013) Independent variations of plant and soil mixtures reveal soil feedback effects on plant community overyielding. J Ecol 101: 287–297. doi: 10.1111/1365-2745.12032
|
| [9] |
Maron JL, Marler M, Klironomos JN, et al. (2011) Soil fungal pathogens and the relationship between plant diversity and productivity. Ecol Lett 14: 36–41. doi: 10.1111/j.1461-0248.2010.01547.x
|
| [10] |
Schnitzer SA, Klironomos JN, Hillerislambers J, et al. (2011) Soil microbes drive the classic plant diversity-productivity pattern. Ecology 92: 296–303. doi: 10.1890/10-0773.1
|
| [11] | Daniels LD, Mariko Y, Werner KL, et al. (2010) Preservation of old-growth forests: A case study of Big Timber Park, Whistler, BC. Can Geogr 52: 367–379. |
| [12] |
Aukema BH, Zhu J, Møller J, et al. (2010) Predisposition to bark beetle attack by root herbivores and associated pathogens: Roles in forest decline, gap formation, and persistence of endemic bark beetle populations. Forest Ecol Manag 259: 374–382. doi: 10.1016/j.foreco.2009.10.032
|
| [13] |
Anagnostakis SL (1987) Chestnut blight: The classical problem of an introduced pathogen. Mycologia 79: 23–37. doi: 10.2307/3807741
|
| [14] | Anagnostakis SL (2002) The effect of multiple importations of pests and pathogens on a native tree. Biol Invasions 3: 245–254. |
| [15] |
Keever C (1953) Present composition of some stands of the former oak-chestnut forest in the southern Blue Ridge Mountains. Ecology 34: 44–54. doi: 10.2307/1930307
|
| [16] |
Anagnostakis SL (2012) Chestnut breeding in the United States for disease and insect resistance. Plant Dis 96: 1392–1403. doi: 10.1094/PDIS-04-12-0350-FE
|
| [17] | Burnham CR (1988) The restoration of the American chestnut. Am Sci 76: 478–486. |
| [18] |
Mccament CL, Mccarthy BC (2005) Two-year response of American chestnut (Castanea dentata) seedlings to shelterwood harvesting and fire in a mixed-oak forest ecosystem. Can J Forest Res 35: 740–749. doi: 10.1139/x05-002
|
| [19] |
Rhoades C, Loftis D, Lewis J, et al. (2009) The influence of silvicultural treatments and site conditions on American chestnut (Castanea dentata) seedling establishment in eastern Kentucky, USA. Forest Ecol Manag 258: 1211–1218. doi: 10.1016/j.foreco.2009.06.014
|
| [20] |
Griscom HP, Griscom BW (2012) Evaluating the ecological niche of American chestnut for optimal hybrid seedling reintroduction sites in the Appalachian ridge and valley province. New Forest 43: 441–455. doi: 10.1007/s11056-011-9291-7
|
| [21] |
Clark S, McNab H, Loftis D, et al. (2012) American chestnut growth and survival five years after planting in two silvicultural treatments in southern Appalachians, USA. Forests 3: 1017–1033. doi: 10.3390/f3041017
|
| [22] |
Clark SL, Schlarbaum SE, Pinchot CC, et al. (2014) Reintroduction of American chestnut in the National Forest System. J Forest 112: 502–512. doi: 10.5849/jof.13-106
|
| [23] |
McCarthy BC, Bauman JM, Keiffer CH (2008) Mine reclamation strategies for the restoration of American chestnut (Castanea dentata). Ecol Res 26: 292–294. doi: 10.3368/er.26.4.292
|
| [24] |
Skousen J, Cook T, Wilson-Kokes L, et al. (2013) Survival and growth of chestnut backcross seed and seedlings on surface mines. J Environ Qual 42: 690. doi: 10.2134/jeq2012.0368
|
| [25] |
Bauman JM, Keiffer CH, McCarthy BC (2014) Backcrossed chestnut seedling performance and blight incidence (Cryphonectria parasitica) in restoration. New Forest 45: 813–828. doi: 10.1007/s11056-014-9439-3
|
| [26] |
Gilland KE, McCarthy BC (2014) Performance and phenotypic variation of American chestnut (Castanea dentata) hybrids on newly reclaimed mine sites in eastern Ohio, USA. Ecol Res 32: 379–387. doi: 10.3368/er.32.4.379
|
| [27] |
Barton C, Miller J, Sena K, et al. (2015) Evaluating the use of tree shelters for direct seeding of Castanea on a surface mine in Appalachia. Forests 6: 3514–3527. doi: 10.3390/f6103514
|
| [28] |
Jacobs DF (2007) Toward development of silvical strategies for forest restoration of American chestnut (Castanea dentata) using blight-resistant hybrids. Biol Conserv 137: 497–506. doi: 10.1016/j.biocon.2007.03.013
|
| [29] |
Palmer JM, Lindner DL, Volk TJ (2008) Ectomycorrhizal characterization of an American chestnut (Castanea dentata)-dominated community in Western Wisconsin. Mycorrhiza 19: 27. doi: 10.1007/s00572-008-0200-7
|
| [30] | Bauman JM, Keiffer CH, Hiremath S (2012) Facilitation of American chestnut (Castanea dentata) seedlings by established Pinus virginiana in mine reclamation. Int J Ecol. |
| [31] |
D'Amico KM, Horton TR, Maynard CA, et al. (2015) Comparison of ectomycorrhizal colonization of transgenic American chestnut with those of the wild type, a conventionally bred hybrid, and related fagaceae species. Appl Environ Microbiol 81: 100–108. doi: 10.1128/AEM.02169-14
|
| [32] | Dulmer KM, Leduc SD, Horton T (2014) Ectomycorrhizal inoculum potential of northeastern US forest soils for American chestnut restoration: Results from field and laboratory bioassays. Mycorrhiza 24: 65–74. |
| [33] |
Jones MD, Hagerman SM, Gillespie M (2002) Ectomycorrhizal colonization and richness of previously colonized, containerized Picea engelmannii does not vary across clearcuts when planted in mechanically site-prepared mounds. Can J Forest Res 32: 1425–1433. doi: 10.1139/x02-069
|
| [34] |
Jasper DA (2007) Beneficial soil microorganisms of the Jarrah Forest and their recovery in bauxite mine restoration in southwestern Australia. Res Ecol 15: S74–S84. doi: 10.1111/j.1526-100X.2007.00295.x
|
| [35] |
Iordache V, Gherghel F, Kothe E (2009) Assessing the effect of disturbance on ectomycorrhiza diversity. Int J Environ Res Pub Health 6: 414–432. doi: 10.3390/ijerph6020414
|
| [36] | Marx DH (1991) The practical significance of ectomycorrhizae in forest establishment. Marcus Wallenberg Foundation Symposia Proceedings 7: 54–90. |
| [37] | Castellano MA (1996) Outplanting performance of mycorrhizal inoculated seedlings, In: Mukerji KG, Editor, Concepts in Mycorrhizal Research, Kluwer Academic Publishers B.V., 223–301. |
| [38] |
Bauman JM, Keiffer CH, Hiremath S, et al. (2013) Soil preparation methods promoting ectomycorrhizal colonization and American chestnut (Castanea dentata) establishment in coal mine restoration. J App Ecol 50: 721–729. doi: 10.1111/1365-2664.12070
|
| [39] | Bauman JM, Adamson J, Brisbin R, et al. (2017) Soil metals and ectomycorrhizal fungi associated with American chestnut hybrids as reclamation trees on formerly coal mined land. Int J Agron. |
| [40] |
Gehring CA, Cobb NS, Whitham TG (1997) Three-way interactions among ectomycorrhizal mutualists, scale insects, and resistant and susceptible pinyon pines. Am Nat 149: 824–841. doi: 10.1086/286026
|
| [41] |
Mueller RCMC, Sthultz CMSM, Martinez TM, et al. (2005) The relationship between stem-galling wasps and mycorrhizal colonization of Quercus turbinella. Can J Bot 83: 1349–1353. doi: 10.1139/b05-105
|
| [42] |
Gehring CA, Whitman TG (1991) Herbivore-driven mycorrhizal mutualism in insect susceptible pinyon pine. Nature 353: 556–557. doi: 10.1038/353556a0
|
| [43] |
Saikkonen KU, Ahonen-Jonnarth U, Markkola AM, et al. (1999) Defoliation and mycorrhizal symbiosis: A functional balance between carbon sources and below-ground sinks. Ecol Lett 2: 19–26. doi: 10.1046/j.1461-0248.1999.21042.x
|
| [44] |
Markkola A, Kuikka K, Rautio P, et al. (2004) Defoliation increases carbon limitation in ectomycorrhizal symbiosis of Betula pubescens. Oecologia 140: 234–240. doi: 10.1007/s00442-004-1587-2
|
| [45] |
Kosola KR, Durall DM, Robertson GP, et al. (2004) Resilience of mycorrhizal fungi on defoliated and fertilized hybrid poplars. Can J Bot 82: 671–680. doi: 10.1139/b04-038
|
| [46] |
Scott PM, Shearer BL, Barber PA, et al. (2013) Relationships between the crown health, fine root and ectomycorrhizae density of declining Eucalyptus gomphocephala. Australas Plant Path 42: 121–131. doi: 10.1007/s13313-012-0152-4
|
| [47] |
Corcobado T, Moreno G, Azul AM, et al. (2015) Seasonal variations of ectomycorrhizal communities in declining Quercus ilex forests: Interactions with topography, tree health status and Phytophthora cinnamomi infections. Forestry 88: 257–266. doi: 10.1093/forestry/cpu056
|
| [48] | Blom JM, Vannini A, Vettraino AM, et al. (2009) Ectomycorrhizal community structure in a healthy and a Phytophthora infected chestnut (Castanea sativa Mill.) stand in central Italy. Mycorrhiza 20: 25–38. |
| [49] |
Kuikka K, Härmä Esa, Markkola A, et al. (2003) Severe defoliation of Scots pine reduces reproductive investment by ectomycorrhizal symbionts. Ecology 84: 2051–2061. doi: 10.1890/02-0359
|
| [50] |
Saravesi K, Markkola A, Rautio P, et al. (2008) Defoliation causes parallel temporal responses in a host tree and its fungal symbionts. Oecologia 156: 117–123. doi: 10.1007/s00442-008-0967-4
|
| [51] |
Altschul SF, Madden TL, Schaffer AA, et al. (1997) Gapped BLAST and PSIBLAST: A new generation of protein database search programs. Nucleic Acids Res 25: 3389–3402. doi: 10.1093/nar/25.17.3389
|
| [52] |
Kãµljalg U, Nilsson RH, Abarenkov K, et al. (2013) Towards a unified paradigm for sequence-based identification of fungi. Mol Ecol 22: 5271–5277. doi: 10.1111/mec.12481
|
| [53] | R Development Core Team R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria, 2009. Available from: http://www.R-project.org. |
| [54] | Oksanen J, Blanchet FG, Kindt R, et al. (2010) Vegan: Community ecology package. Available from: http://CRAN.R-project.org/package=vegan. |
| [55] | Luoma DL, Eberhart JL (2014) Relationships between Swiss needle cast and ectomycorrhizal fungus diversity. Mycorrhiza 4: 666–675. |
| [56] | Gehring CA, Whitman TG (2002) Mycorrhizae-herbivore interactions: population and community consequences, In: van der Heiden MGA, IR Sanders, Editors, Mycorrhizal Ecology, Berlin: Springer, 295–320. |
| [57] |
Horton BM, Glen M, Davidson NJ, et al. (2013) Temperate eucalypt forest decline is linked to altered ectomycorrhizal communities mediated by soil chemistry. Forest Ecol Manag 302: 329–337. doi: 10.1016/j.foreco.2013.04.006
|
| [58] |
Agerer R (2001) Exploration types of ectomycorrhizae. Mycorrhiza 11: 107–114. doi: 10.1007/s005720100108
|
| [59] |
Peay KG, Kennedy PG, Bruns TD (2011) Rethinking ectomycorrhizal succession: Are root density and hyphal exploration types drivers of spatial and temporal zonation? Fungal Ecol 4: 233–240. doi: 10.1016/j.funeco.2010.09.010
|
| [60] |
Cairney JWG (1992) Translocation of solutes in ectomycorrhizal and saprotrophic rhizomorphs. Mycol Res 96: 135–141. doi: 10.1016/S0953-7562(09)80928-3
|
| [61] |
Agerer R (2006) Fungal relationships with structural identity of their ectomycorrhizae. Mycol Prog 5: 67–107. doi: 10.1007/s11557-006-0505-x
|
| [62] | Hobbie EA, Agerer R (2010) Nitrogen isotops in ectomycorrhizal sporocarps correspond to belowground exploration types. Plant Soil 32: 71–83. |
| [63] |
Lilleskov EA, Hobbie EA, Horton TR (2011) Conservation of ectomycorrhizal fungi: Exploring the linkages between functional and taxonomic responses to anthropogenic N deposition. Fungal Ecol 4: 174–183. doi: 10.1016/j.funeco.2010.09.008
|
| [64] | Ericsson T (1995) Growth and shoot: Root ratio of seedlings in relation to nutrient availability. Plant Soil 169: 205–214. |
| [65] |
Bödeker IT, Nygren CM, Taylor AF, et al. (2009) ClassII peroxidase-encoding genes are present in a phylogenetically wide range of ectomycorrhizal fungi. ISME J 3: 1387–1395. doi: 10.1038/ismej.2009.77
|
| [66] |
Kranabetterj M (2014) Ectomycorrhizal fungi and the nitrogen economy of conifers-implications for genecology and climate change mitigation. Botany 92: 417–423. doi: 10.1139/cjb-2013-0198
|
| [67] |
Koide RT, Fernandez CW, Malcolm C (2014) Determining place and process: Functional traits of ectomycorrhizal fungi that affect both community structure and ecosystem function. New Phytol 201: 433–439. doi: 10.1111/nph.12538
|
| [68] |
Clemmensen KE, Bahr A, Ovaskainen O, et al. (2013) Roots and associated fungi drive long term carbon sequestration in boreal forest. Science 339: 1615–1618. doi: 10.1126/science.1231923
|
| [69] | Montecchio L, Causin R, Rossi S (2004) Changes in ectomycorrhizal diversity in a declining Quercus ilex coastal forest. Phytopathol Mediterr 43: 26–34. |
| [70] |
Dickie IA, Schnitzer SA, Reich PB (2005) Spatially disjunct effects of co-occurring competition and facilitation. Ecol Lett 8: 1191–1200. doi: 10.1111/j.1461-0248.2005.00822.x
|
| [71] | Pigott C (2006) Survival of mycorrhiza formed by Cenococcum geophilum FR. in dry soils. New Phytol 92: 513–517. |
| [72] | Bell JE, Leeper RD, Palecki MA, et al. (2015) Evaluation of the 2012 drought with a newly established national soil monitoring network. Vadose Zone J 14: 148. |
| [73] |
Fernandez CW, Mccormack ML, Hill JM, et al. (2013) On the persistence of Cenococcum geophilum mycorrhizas and its implications for forest carbon and nutrient cycles. Soil Biol Biochem 65: 141–143. doi: 10.1016/j.soilbio.2013.05.022
|
| [74] |
Singaravelan N, Grishkan I, Beharav A, et al. (2008) Adaptive melanin response of the soil fungus Aspergillus niger to UV radiation stress at "Evolution Canyon", Mount Carmel, Israel. PLoS One 3: e2993. doi: 10.1371/journal.pone.0002993
|
| [75] |
Gadd GM, Derome L (1988) Biosorption of copper by fungal melanin. Appl Microbiol Biotechnol 29: 610–617. doi: 10.1007/BF00260993
|
| [76] |
Kogej T, Gorbushina AA, Gunde-Cimerman N (2006) Hypersaline conditions induce changes in cell-wall melanization and colony structure in a halophilic and a xerophilic black. Mycol Res 110: 713–724. doi: 10.1016/j.mycres.2006.01.014
|
| [77] | Bauman JM, Keiffer CH, Hiremath S (2011) The influence of inoculated and native ectomycorrhizal fungi on morphology, physiology and survival of American chestnut, In: Barnhisel RI, Editor, The American Society of Mining and Reclamation Proceedings. Sciences Leading to Success, Lexington, KY, 16–37. |
| [78] |
Kennedy P, Bruns TD (2005) Priority effects determine the outcome of ectomycorrhizal competition between two Rhizopogon species colonizing Pinus muricata seedlings. New Phytol 166: 631–638. doi: 10.1111/j.1469-8137.2005.01355.x
|
| [79] | Braun EL (1950) Deciduous forests of Eastern North America. New York: McGraw-Hill Book Co. Inc., 596. |
| [80] |
Talbot JM, Bruns TD, Smith DP, et al. (2013) Independent roles of ectomycorrhizal and saprotrophic communities in soil organic matter decomposition. Soil Biol Biochem 57: 282–291. doi: 10.1016/j.soilbio.2012.10.004
|
| [81] |
Li A, Fahey TJ, Pawlowska TE, et al. (2015) Fine root decomposition, nutrient mobilization an fungal communities in a pine forest ecosystem. Soil Biol Biochem 83: 76–83. doi: 10.1016/j.soilbio.2015.01.019
|
| [82] |
Fernandez CW, Koide RT (2013) The function of melanin in the ectomycorrhizal fungus Cenococcum geophilum under water stress. Fungal Ecol 6: 479–486. doi: 10.1016/j.funeco.2013.08.004
|
| [83] |
Bever JD (2002) Host-specificity of AM fungal population growth rates can generate feedback on plant growth. Plant Soil 244: 281–290. doi: 10.1023/A:1020221609080
|
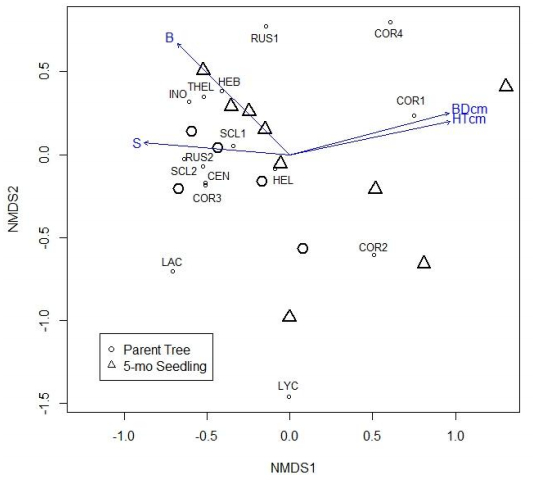
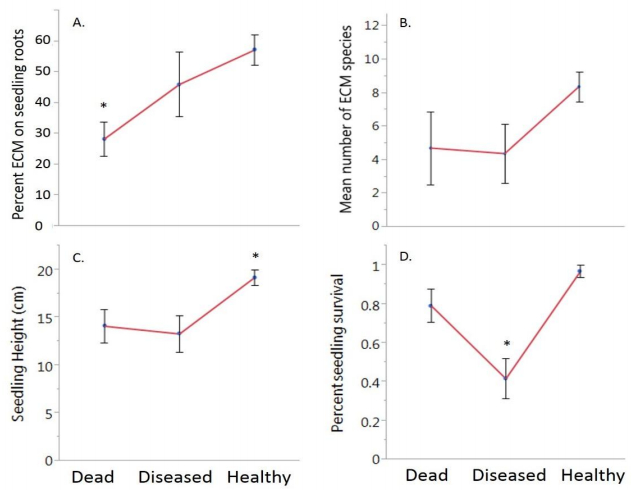
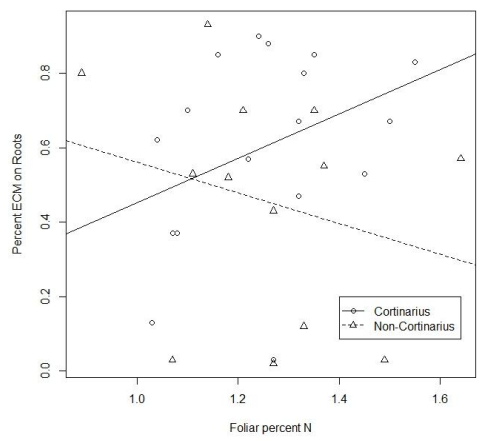
Figures(3) / Tables(2)

Jenise M. Bauman, Sarah Francino, Amy Santas. Interactions between ectomycorrhizal fungi and chestnut blight (Cryphonectria parasitica) on American chestnut (Castanea dentata) used in coal mine restoration[J]. AIMS Microbiology, 2018, 4(1): 104-122. doi: 10.3934/microbiol.2018.1.104







 DownLoad:
DownLoad: