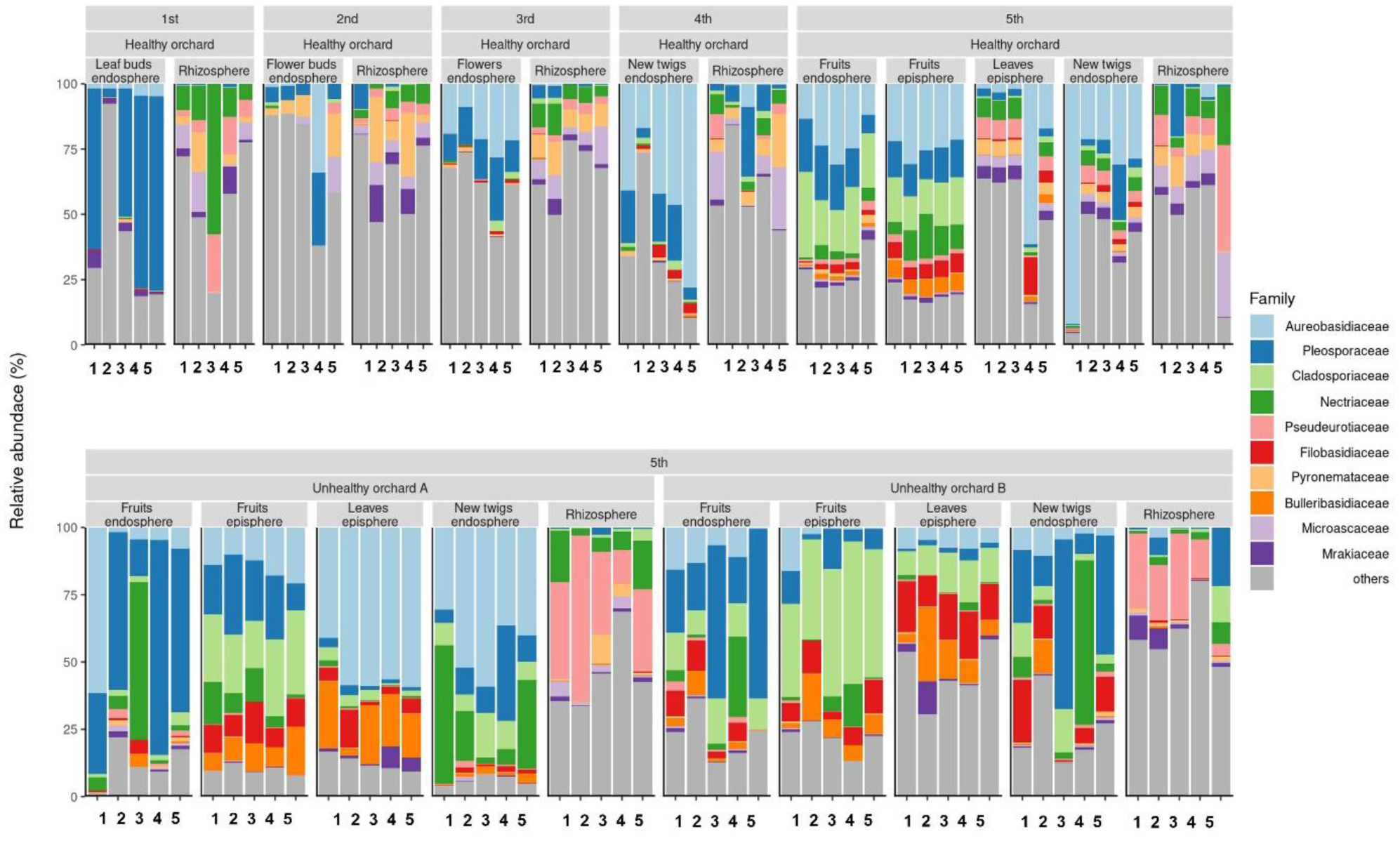
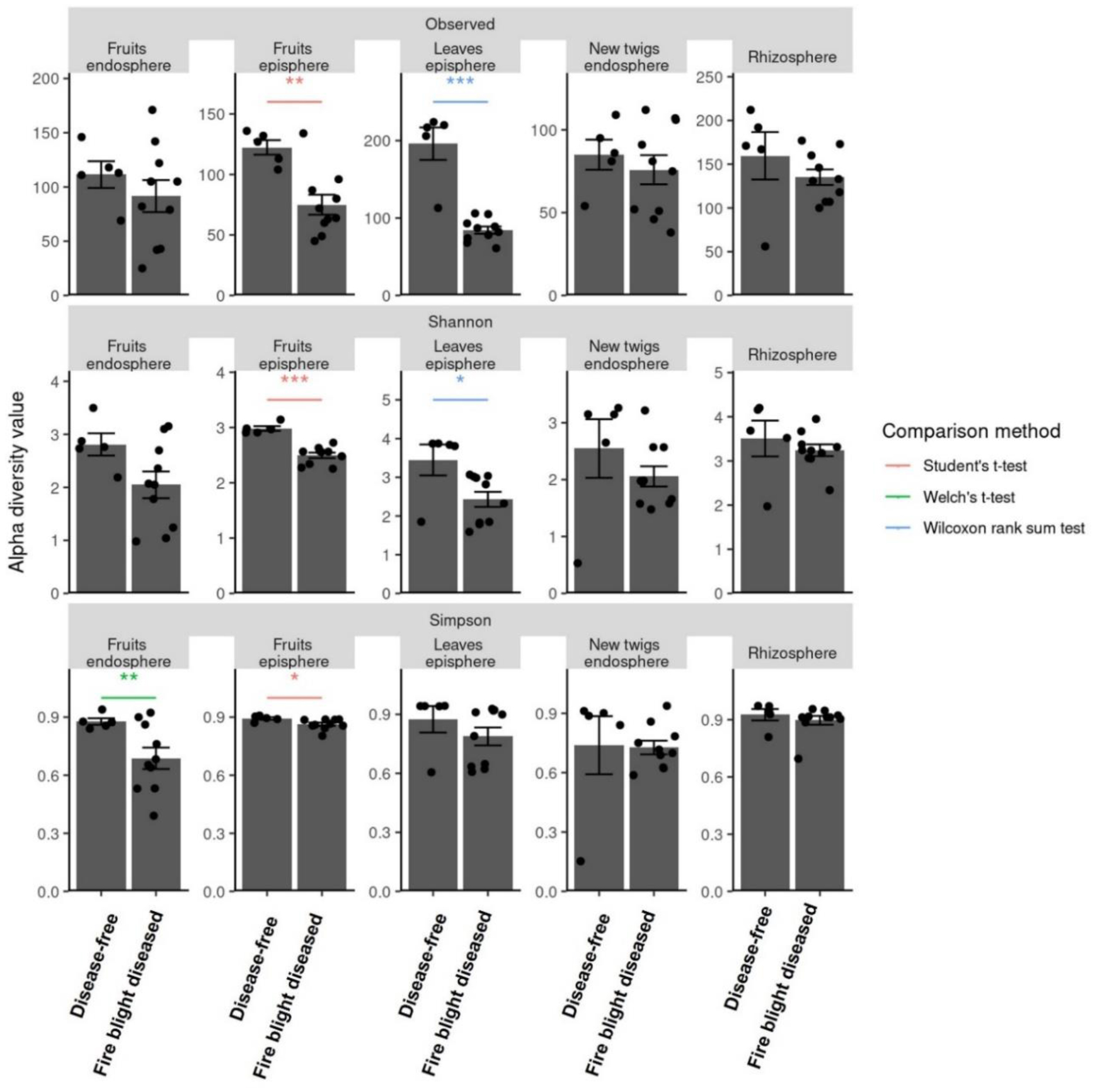
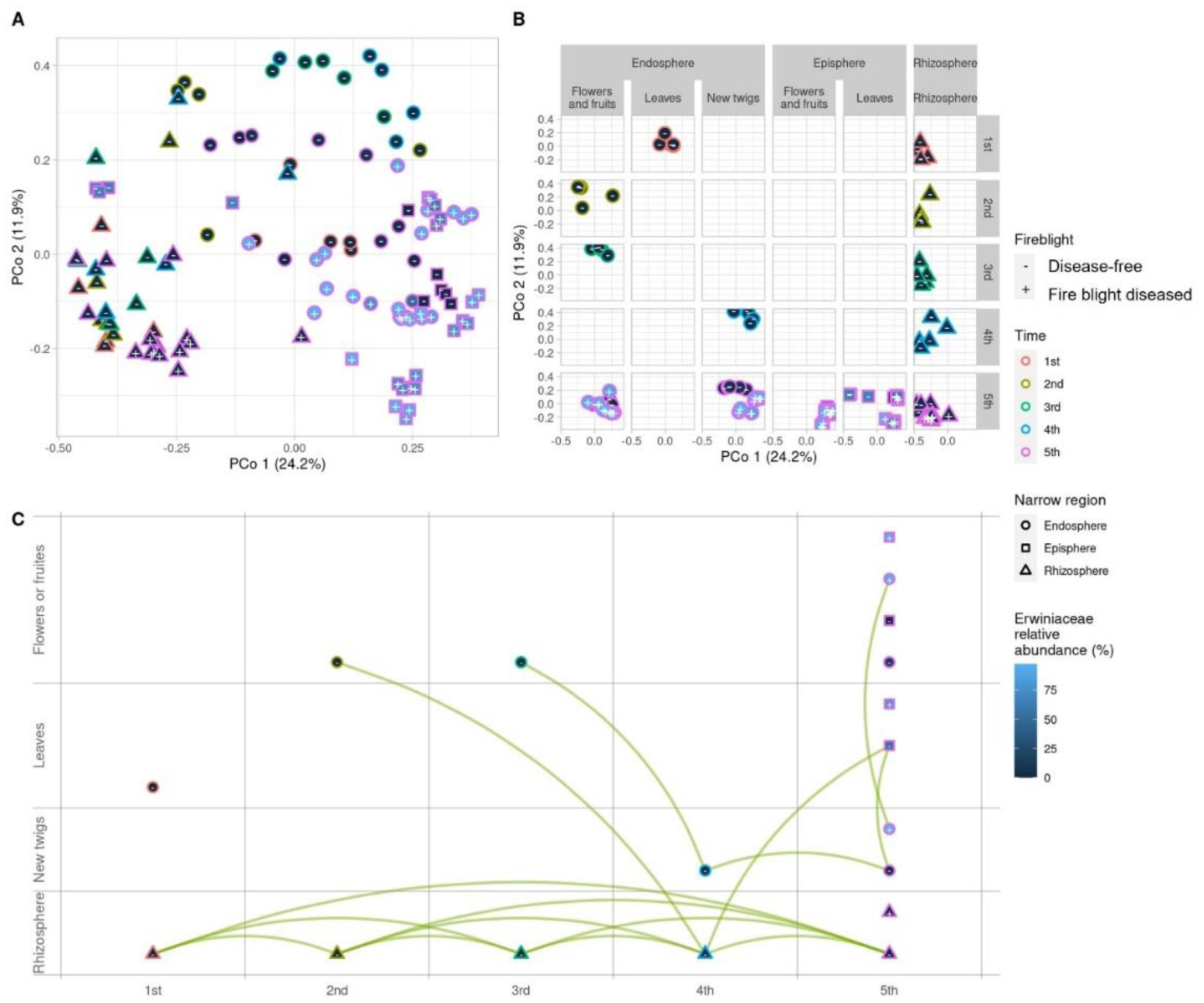
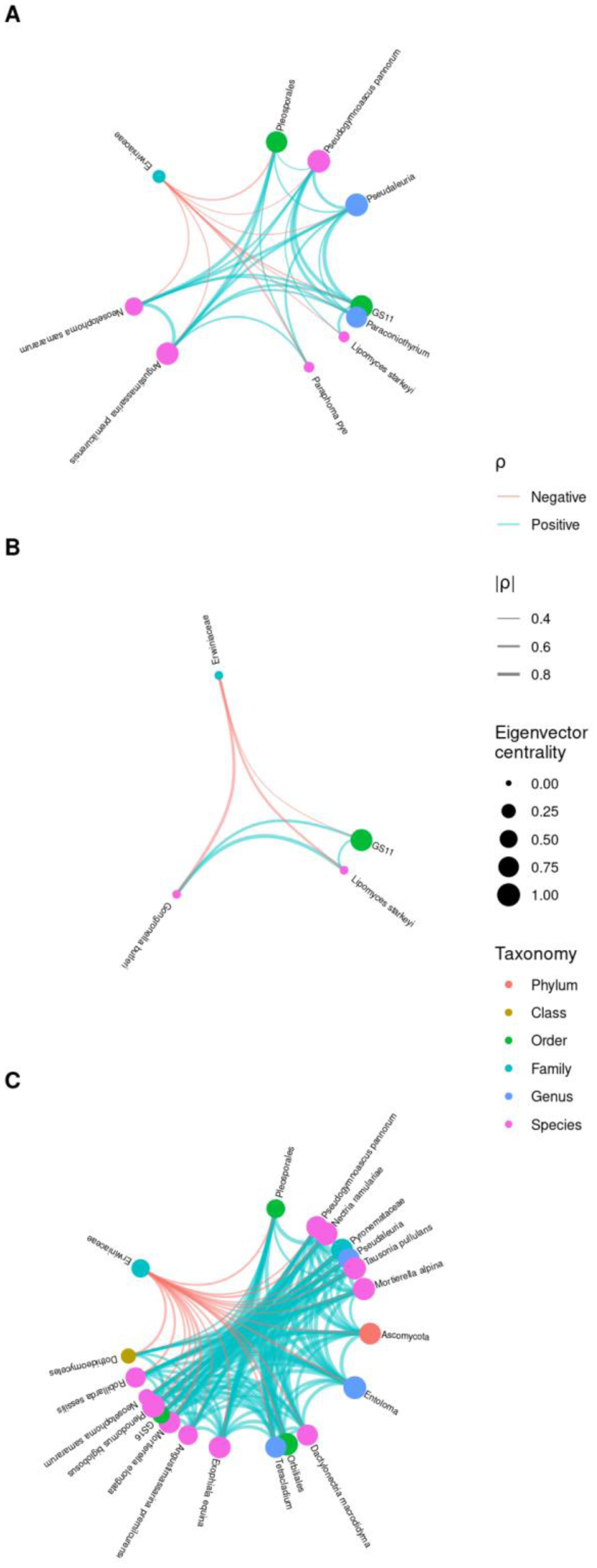
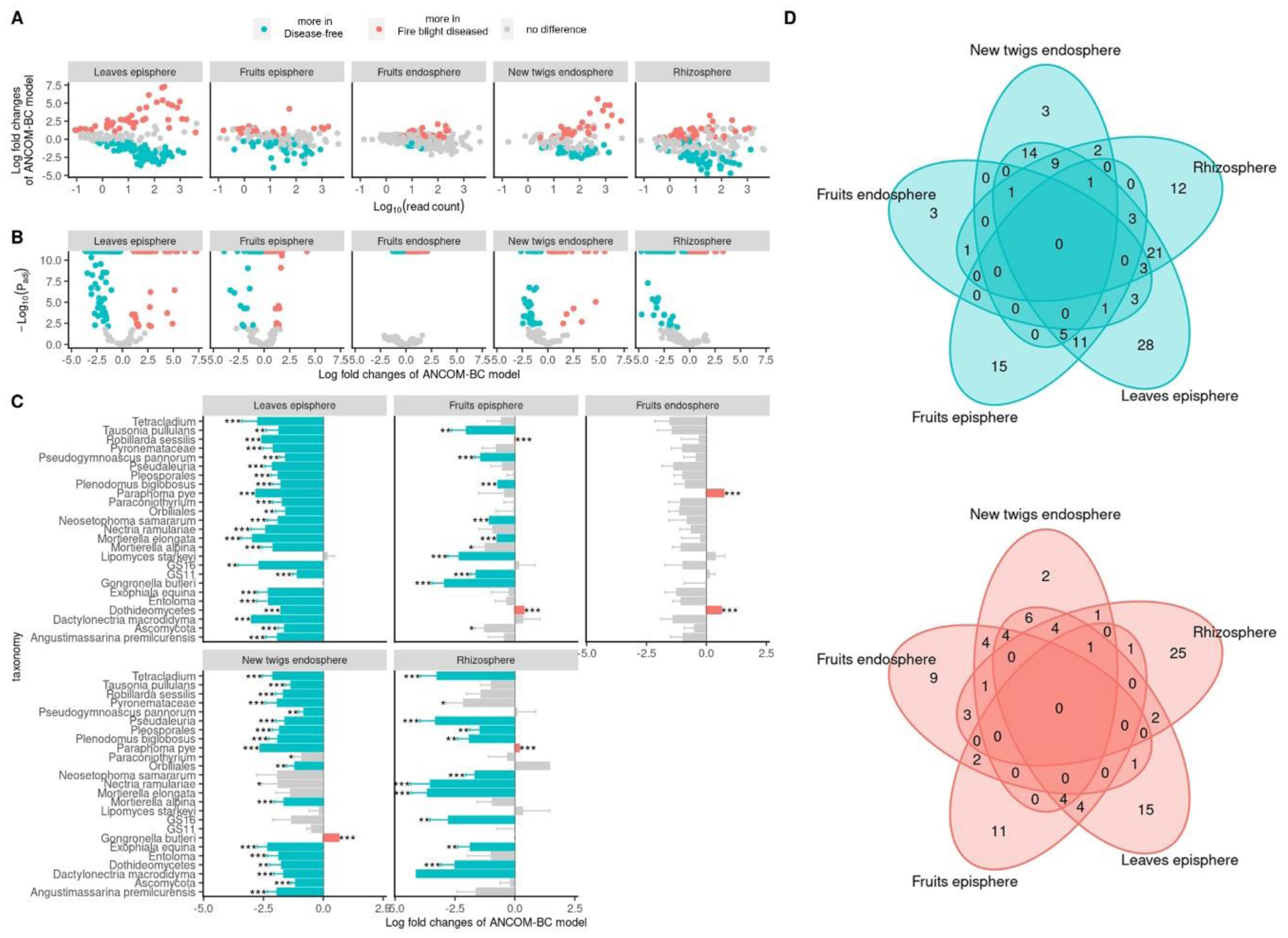
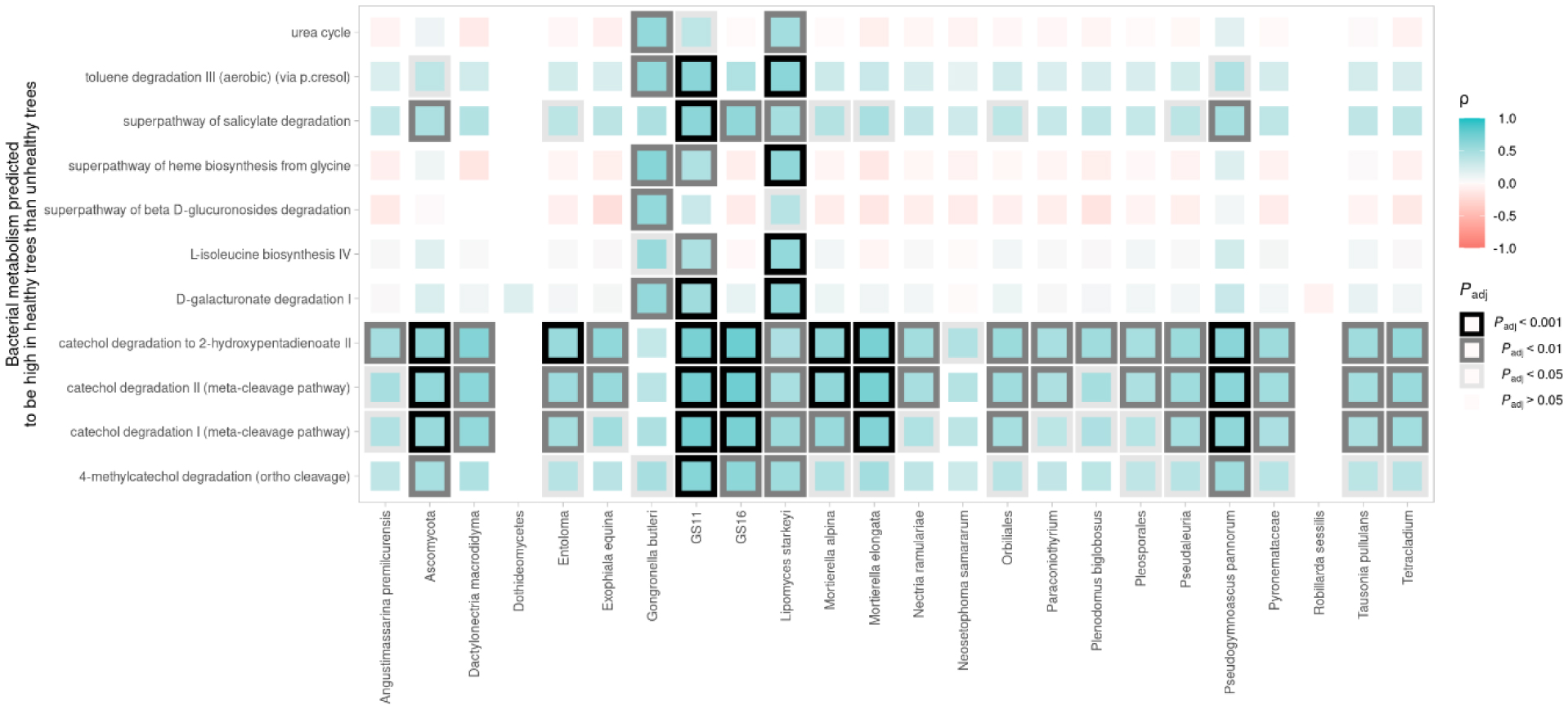
Fire blight disease, caused by the bacterial pathogen Erwinia amylovora, has been a significant concern for over 50 countries worldwide. The efficacy of chemical pesticides currently available for disease control is limited. To address this issue, research is being conducted to explore environmentally friendly control methods, particularly biological control using beneficial microorganisms. However, there is limited research on the apple microbiota community and minimal research has been conducted on fungal communities that may exhibit reliable performance in apple trees. Therefore, our objective was to analyze the fungal communities present in apples at different developmental stages and in different tissues, aiming to identify potential biological control agents for fire blight disease. Our findings indicate that the fungal communities present in apple buds, flowers and leaves play an important role in inhibiting the invasion of E. amylovora. Specifically, we propose GS11 and Lipomyces starkeyi as potential keystone taxa that respond to fire blight disease. These findings provide insights into the continuity and discontinuity of fungal community structure in different developmental stages of apples and offer predictions for potential biological control agents for fire blight disease.
Citation: Su In Lee, Gyeongjun Cho, Su-Hyeon Kim, Da-Ran Kim, Youn-Sig Kwak. Mycobiota community and fungal species response to development stage and fire blight disease in apples[J]. AIMS Microbiology, 2023, 9(3): 554-569. doi: 10.3934/microbiol.2023029
Fire blight disease, caused by the bacterial pathogen Erwinia amylovora, has been a significant concern for over 50 countries worldwide. The efficacy of chemical pesticides currently available for disease control is limited. To address this issue, research is being conducted to explore environmentally friendly control methods, particularly biological control using beneficial microorganisms. However, there is limited research on the apple microbiota community and minimal research has been conducted on fungal communities that may exhibit reliable performance in apple trees. Therefore, our objective was to analyze the fungal communities present in apples at different developmental stages and in different tissues, aiming to identify potential biological control agents for fire blight disease. Our findings indicate that the fungal communities present in apple buds, flowers and leaves play an important role in inhibiting the invasion of E. amylovora. Specifically, we propose GS11 and Lipomyces starkeyi as potential keystone taxa that respond to fire blight disease. These findings provide insights into the continuity and discontinuity of fungal community structure in different developmental stages of apples and offer predictions for potential biological control agents for fire blight disease.

| [1] | O'Rourke D (2021) Economic importance of the World apple industry. The apple genome . Switzerland: Springer 1-18. |
| [2] |
Wassermann B, Müller H, Berg G (2019) An apple a day: which bacteria do we eat with organic and conventional apples?. Front Microbiol 10: 1629. https://doi.org/10.3389/fmicb.2019.01629 
|
| [3] | Sutton TB, Aldwinckle SH, Agnello AM, et al. (2014) Compendium of apple and pear diseases and pest. APS publication. |
| [4] |
Thapa R, Zhang K, Snavely N, et al. (2020) The plant pathology challenge 2020 data set to classify foliar disease in apples. Appl Plant Sci 8: e11390. https://doi.org/10.1002/aps3.11390
|
| [5] |
Sheikh A, Ahmad BM, Ahmad BZ (2019) Validation of fungicide spray schedule of conventional apple orchards under high density apple orchard system against scab an Alternaria leaf blotch. Pestic Res J 31: 92-95. https://doi.org/10.5958/2249-524X.2019.00017.7
|
| [6] | van der Zwet T (1996) Present worldwide distribution of fire blight. ISHA Acta Horticul 411. https://doi.org/10.17660/ActaHortic.1996.411.2 |
| [7] |
Myung I, Lee J, Yun M, et al. (2016) Fire blight of apple, caused by Erwinia amylovora, a new disease in Korea. Plant Dis 100: 1774. https://doi.org/10.1094/PDIS-01-16-0024-PDN
|
| [8] |
Oh C, Beer SV (2005) Molecular genetics of Erwinia amylovora involved in the development of fire blight. FEMS Microbiol Lett 253: 185-192. https://doi.org/10.1016/j.femsle.2005.09.051
|
| [9] |
Bayot RG, Ries SM (1986) Role of motility in apple blossom infection by Erwinia amylovora and studies of fire blight control with attractant and repellent compounds. Phytopathology 76: 441-445. https://doi.org/10.1094/PHYTO-76-441
|
| [10] |
Chatterjee A (2001) Fire blight: The disease and its causative agent, Erwinia amylovora?. Eur J Plant Pathol 107: 569.
|
| [11] |
Choi JH, Kim J, Park DH (2022) Evidence of greater competitive fitness of Erwinia amylovora over E. pyrifoliae in Korean isolates. Plant Pathol J 38: 355-365. https://doi.org/10.5423/PPJ.OA.04.2022.0056
|
| [12] |
Ham H, Oh G, Park DS, et al. (2022) Survey of oxolinic acid-resistant Erwinia amylovora in Korean apple and pear orchards, and the fitness impact of constructed mutants. Plant Pathol J 38: 482-489. https://doi.org/10.5423/PPJ.OA.04.2022.0059
|
| [13] |
Slack SM, Walters KJ, Outwater CA, et al. (2021) Effect of Kausgamycin, Oxytetracycline, and Streptomycin on in orchard population dynamics of Erwinia amylovora on apple flower stigmas. Plant Dis 105: 1843-1850. https://doi.org/10.1094/PDIS-07-20-1469-RE
|
| [14] |
Förster H, McGhee GC, Sundin GW, et al. (2015) Characterization of Streptomycin resistance in isolates of Erwinia amylovora in California. Phytopathology 105: 1302-1310. https://doi.org/10.1094/PHYTO-03-15-0078-R
|
| [15] | Madrid AJ, Ivey ML (2022) An overview of Streptomycin resistance Erwinia amylovora from Ohio apple orchards. Plant Health Prog 24. https://doi.org/10.1094/PHP-08-22-0074-RS |
| [16] |
Weller DM, Cook RJ (1983) Suppression of take-all of wheat by seed treatment with fluorescent Pseudomonads. Phytopathology 73: 463-469. https://doi.org/10.1094/phyto-73-463
|
| [17] |
Zhang J, Mavrodi DV, Yang M, et al. (2020) Pseudomonas synxantha 2-79 transformed with pyrrolnitin biosynthesis genes has improved biocontrol activity against soilborne pathogens of wheat and canola. Phytopathology 110: 1010-1017. https://doi.org/10.1094/PHYTO-09-19-0367-R
|
| [18] |
Dagher F, Nickzar A, Zheng J, et al. (2021) Characterization of the biocontrol activity of three bacterial isolates against the phytopathogen Erwinia amylovora. MicrobiologyOpen 10: e1202. https://doi.org/10.1002/mbo3.1202
|
| [19] |
Medhioub I, Cheffi M, Tounsi S, et al. (2022) Study of Bacillus velezensis OEE1 potentialities in the biocontrol against Erwinia amylovora, causal agent of fire blight disease of rosaceous plants. Biol Control 167: 1084842. https://doi.org/10.1016/j.biocontrol.2022.104842
|
| [20] |
Temple TN, Thompson EC, Uppala S, et al. (2020) Floral colonization dynamics and specificity of Aureobasidium pullulans strains used to suppress fire blight of pome fruit. Plant Dis 104: 121-128. https://doi.org/10.1094/PDIS-09-18-1512-RE
|
| [21] |
Kim D, Cho G, Jeon C, et al. (2019) A mutualistic interaction between Streptomyces bacteria, strawberry plants and pollinating bees. Nat Commun 10: 4802. https://doi.org/10.1038/s41467-019-12785-3
|
| [22] |
Kim S, Cho G, Lee SI, et al. (2021) Comparison of bacterial community of healthy and Erwinia amylovora infected apples. Plant Pathol J 37: 396-403. https://doi.org/10.5423/PPJ.NT.04.2021.0062
|
| [23] |
Kong HG, Ham H, Lee M, et al. (2021) Microbial community dysbiosis and functional gene content changes in apple flowers due to fire blight. Plant Pathol J 37: 404-412. https://doi.org/10.5423/PPJ.NT.05.2021.0072
|
| [24] |
Francesco AD, Foggia MD, Corbetta M, et al. (2021) Biocontrol activity and plant growth promotion exerted by Aureobasidium pullulans strains. J Plant Growth Reg 40: 1233-1244. https://doi.org/10.1007/s00344-020-10184-3
|
| [25] |
Francesco AD, Zajc J, Stenberg JA (2023) Aureobasidium spp.: Diversity, versatility, and agricultural utility. Horticulturae 9: 59. https://doi.org/10.3390/horticulturae9010059
|
| [26] |
Rusin C, Francesco AD, Foggia MD, et al. (2019) An emerging problem affecting apple production: Neofusicoccum parvum, Aureobasidium pullulans L1 and L8 strains as an alternative control strategy. Biol Control 134: 157-162. https://doi.org/10.1016/j.biocontrol.2019.04.015
|
| [27] |
Dai Z, Deng S, Culley DE, et al. (2017) Agrobacterium tumefaciens-mediated transformation of oleaginous yeast Lipomyces species. Appl Microbiol Biotechnol 101: 6099-6110. https://doi.org/10.1007/s00253-017-8357-7
|
| [28] |
Mukherjee A, Verma JP, Gaurav AK, et al. (2020) Yeast a potential bio-agent: Future for plant growth and postharvest disease management for sustainable agriculture. Appl Microbiol Biotechnol 104: 1497-1510. https://doi.org/10.1007/s00253-019-10321-3
|
 microbiol-09-03-029-s001.pdf microbiol-09-03-029-s001.pdf |
 |
Figures(6) / Tables(2)

Su In Lee, Gyeongjun Cho, Su-Hyeon Kim, Da-Ran Kim, Youn-Sig Kwak. Mycobiota community and fungal species response to development stage and fire blight disease in apples[J]. AIMS Microbiology, 2023, 9(3): 554-569. doi: 10.3934/microbiol.2023029







 DownLoad:
DownLoad: