Citation: Sébastien Bernacchi, Alexander Krajete, Christoph Herwig. Experimental workflow for developing a feed forward strategy to control biomass growth and exploit maximum specific methane productivity of Methanothermobacter marburgensis in a biological methane production process (BMPP)[J]. AIMS Microbiology, 2016, 2(3): 262-277. doi: 10.3934/microbiol.2016.3.262

| [1] |
Demirbas MF, Balat M (2006) Recent advances on the production and utilization trends of bio-fuels: A global perspective. Energy Convers Manag 47: 2371–2381. doi: 10.1016/j.enconman.2005.11.014

|
| [2] |
Demirbas A (2007) Progress and recent trends in biofuels. Prog Energy Combust Sci 33: 1–18. doi: 10.1016/j.pecs.2006.06.001
|
| [3] |
Pinto AC, Guarieiro LLN, Rezende MJC, et al. (2005) Biodiesel: an overview. J Braz Chem Soc 16: 1313–1330. doi: 10.1590/S0103-50532005000800003
|
| [4] | Jajesniak P, Omar Ali HEM, Wong TS (2014) Carbon Dioxide Capture and Utilization using Biological Systems: Opportunities and Challenges. J Bioprocess Biotech 4:155. |
| [5] | Aresta M, Dibenedetto A, Angelini A (2014) Catalysis for the Valorization of Exhaust Carbon: from CO2 to Chemicals, Materials, and Fuels. Technological Use of CO2. Chem Rev 114: 1709–1742. |
| [6] |
Abubackar HN, Veiga MC, Kennes C (2011) Biological conversion of carbon monoxide: rich syngas or waste gases to bioethanol. Biofuels Bioprod Biorefining 5: 93–114. doi: 10.1002/bbb.256
|
| [7] |
Li H, Liao JC (2013) Biological conversion of carbon dioxide to photosynthetic fuels and electrofuels. Energy Environ Sci 6: 2892–2899. doi: 10.1039/c3ee41847b
|
| [8] |
Demirbas A (2008) Biofuels sources, biofuel policy, biofuel economy and global biofuel projections. Energy Convers Manag 49: 2106–2116. doi: 10.1016/j.enconman.2008.02.020
|
| [9] | Bernacchi S, Lorantfy B, Martinez E, et al. (2014) Restructuring renewable energy sources for more efficient biofuels production with extremophilic microorganisms. Symposium Energieinnovation, Graz, Austria. |
| [10] |
Blanch HW (2012) Bioprocessing for biofuels. Curr Opin Biotechnol 23: 390–395. doi: 10.1016/j.copbio.2011.10.002
|
| [11] |
Ghimire A, Frunzo L, Pirozzi F, et al. (2015) A review on dark fermentative biohydrogen production from organic biomass: Process parameters and use of by-products. Appl Energy 144: 73–95. doi: 10.1016/j.apenergy.2015.01.045
|
| [12] |
Rittmann SK-MR, Lee HS, Lim JK, et al. (2015) One-carbon substrate-based biohydrogen production: Microbes, mechanism, and productivity. Biotechnol Adv 33: 165–177. doi: 10.1016/j.biotechadv.2014.11.004
|
| [13] | Patni N, Shah P, Agarwal S, et al. (2013) Alternate Strategies for Conversion of Waste Plastic to Fuels. ISRN Renew Energy 2013: 1–7. |
| [14] |
Rodríguez Couto S (2008) Exploitation of biological wastes for the production of value‐added products under solid‐state fermentation conditions. Biotechnol J 3: 859–870. doi: 10.1002/biot.200800031
|
| [15] |
Angenent LT, Karim K, Al-Dahhan MH, et al. (2004) Production of bioenergy and biochemicals from industrial and agricultural wastewater. Trends Biotechnol 22: 477–485. doi: 10.1016/j.tibtech.2004.07.001
|
| [16] |
van Groenestijn JW, Kraakman NJR (2005) Recent developments in biological waste gas purification in Europe. Chem Eng J 113: 85–91. doi: 10.1016/j.cej.2005.03.007
|
| [17] |
Kennes C, Rene ER, Veiga MC (2009) Bioprocesses for air pollution control. J Chem Technol Biotechnol 84: 1419–1436. doi: 10.1002/jctb.2216
|
| [18] |
Choi D, Chipman DC, Bents SC, et al. (2010) A Techno-economic Analysis of Polyhydroxyalkanoate and Hydrogen Production from Syngas Fermentation of Gasified Biomass. Appl Biochem Biotechnol 160: 1032–1046. doi: 10.1007/s12010-009-8560-9
|
| [19] | Rittmann S, Seifert A, Herwig C (2013) Essential prerequisites for successful bioprocess development of biological CH4 production from CO2 and H2. Crit Rev Biotechnol 1–12. |
| [20] |
Yasin M, Jeong Y, Park S, et al. (2015) Microbial synthesis gas utilization and ways to resolve kinetic and mass-transfer limitations. Bioresour Technol 177: 361–374. doi: 10.1016/j.biortech.2014.11.022
|
| [21] |
Porqueras EM, Rittmann S, Herwig C (2012) Biofuels and CO 2 neutrality: an opportunity. Biofuels 3: 413–426. doi: 10.4155/bfs.12.25
|
| [22] |
Bernacchi S, Weissgram M, Wukovits W, et al. (2014) Process efficiency simulation for key process parameters in biological methanogenesis. AIMS Bioeng 1: 53–71. doi: 10.3934/bioeng.2014.1.53
|
| [23] | Lehner M, Tichler R, Steinmüller H, et al. (2014) Power-to-Gas: Technology and Business Models. Springer International Publishing. |
| [24] | Götz M, Koch AM, Graf F (2014) State of the Art and Perspectives of CO2 Methanation Process Concepts for Power-to-Gas Applications. International Gas Union Research Conference, Copenhagen. |
| [25] |
Seifert AH, Rittmann S, Bernacchi S, et al. (2013) Method for assessing the impact of emission gasses on physiology and productivity in biological methanogenesis. Bioresour Technol 136: 747–751. doi: 10.1016/j.biortech.2013.03.119
|
| [26] |
Götz M, Lefebvre J, Mörs F, et al. (2016) Renewable Power-to-Gas: A technological and economic review. Renew Energy 85: 1371–1390. doi: 10.1016/j.renene.2015.07.066
|
| [27] | Martin MR, Fornero JJ, Stark R, et al. (2013) A Single-Culture Bioprocess of Methanothermobacter thermautotrophicus to Upgrade Digester Biogas by CO2-to-CH4 Conversion with H2. Archaea 2013: 1–11. |
| [28] |
Sambusiti C, Bellucci M, Zabaniotou A, et al. (2015) Algae as promising feedstocks for fermentative biohydrogen production according to a biorefinery approach: A comprehensive review. Renew. Sustain Energy Rev 44: 20–36. doi: 10.1016/j.rser.2014.12.013
|
| [29] | Cusick RJ (1974) Space station prototype Sabatier reactor design verification testing. presented at the Intersociety Conference on Environmental Systems, Seattle, WA, US. |
| [30] | Murdoch K, Goldblatt L, Carrasquillo R, et al. (2015) Sabatier Methanation Reactor for Space Exploration. |
| [31] | Bernacchi S, Seifert AH, Rittmann S (2013) Benefits of Biological Methanation. presented at the conference: DBI-Fachforum Energiespeicher - Pilotprojekte, Berlin, Germany. |
| [32] |
Li J, Wong C-F, Wong MT, et al. (2014) Modularized Evolution in Archaeal Methanogens Phylogenetic Forest. Genome Biol Evol 6: 3344–3359. doi: 10.1093/gbe/evu259
|
| [33] |
Rittmann S, Seifert A, Herwig C (2012) Quantitative analysis of media dilution rate effects on Methanothermobacter marburgensis grown in continuous culture on H2 and CO2. Biomass Bioenergy 36: 293–301. doi: 10.1016/j.biombioe.2011.10.038
|
| [34] |
Seifert AH, Rittmann S, Herwig C (2014) Analysis of process related factors to increase volumetric productivity and quality of biomethane with Methanothermobacter marburgensis. Appl Energy 132: 155–162. doi: 10.1016/j.apenergy.2014.07.002
|
| [35] |
Bernacchi S, Rittmann S, Seifert AH, et al. (2014) Experimental methods for screening parameters influencing the growth to product yield (Y(x/CH4)) of a biological methane production (BMP) process performed with Methanothermobacter marburgensis. AIMS Bioeng 1: 72–86. doi: 10.3934/bioeng.2014.2.72
|
| [36] | Schill N, van Gulik WM, Voisard D, et al. (1996) Continuous cultures limited by a gaseous substrate: Development of a simple, unstructured mathematical model and experimental verification with Methanobacterium thermoautotrophicum. Biotechnol Bioeng 51: 645–658. |
| [37] | Archer DB (1985) Uncoupling of Methanogenesis from Growth of Methanosarcina barkeri by Phosphate Limitation. Appl Environ Microbiol 50: 1233–1237. |
| [38] | Wrede C, Walbaum U, Ducki A, et al (2913) Localization of Methyl-Coenzyme M Reductase as Metabolic Marker for Diverse Methanogenic Archaea. Archaea 2013: 1–7. |
| [39] | Bernacchi S, Seifert AH, Krajete A, et al. (2013) Method and system for producing methane using methanogenic microorganisms and applying specific nitrogen concentrations in the liquid phase. Available from: http://www.google.com/patents/WO2014128300A1?cl=de. |
| [40] |
Schönheit P, Moll J, Thauer RK (1980) Growth parameters (K s, μmax, Y s) of Methanobacterium thermoautotrophicum. Arch Microbiol 127: 59–65. doi: 10.1007/BF00414356
|
| [41] |
Fuchs G, Stupperich E, Thauer RK (1978) Acetate assimilation and the synthesis of alanine, aspartate and glutamate in Methanobacterium thermoautotrophicum. Arch Microbiol 117: 61–66. doi: 10.1007/BF00689352
|
| [42] |
Thauer RK, Kaster A-K, Goenrich M (2010) Hydrogenases from Methanogenic Archaea, Nickel, a Novel Cofactor, and H 2 Storage. Annu Rev Biochem 79: 507–536. doi: 10.1146/annurev.biochem.030508.152103
|
| [43] |
Duin E, Cosper N, Mahlert F (2003) Coordination and geometry of the nickel atom in active methyl-coenzyme M reductase from Methanothermobacter marburgensis as detected by X-ray absorption spectroscopy. J Biol Inorg Chem 8: 141–148. doi: 10.1007/s00775-002-0399-2
|
| [44] |
Ullmann E, Tan TC, Gundinger T, et al. (2014) A novel cytosolic NADH:quinone oxidoreductase from Methanothermobacter marburgensis. Biosci Rep 34: 893–904. doi: 10.1042/BSR20140143
|
| [45] | Martin DD, Ciulla RA, Roberts MF (1999) Osmoadaptation in Archaea. Appl Environ Microbiol 65: 1815–1825. |
| [46] | Ciulla R, Clougherty C, Belay N (1994) Halotolerance of Methanobacterium thermoautotrophicum delta H and Marburg. J Bacteriol 176: 3177–3187. |
| [47] | Kempf B, Bremer E (1988) Stress responses ofBacillus subtilis to high osmolarity environments: Uptake and synthesis of osmoprotectants. J Biosci 23: 447–455. |
| [48] |
de Poorter LMI, Geerts WJ, Keltjens JT (2007) Coupling of Methanothermobacter thermautotrophicus Methane Formation and Growth in Fed-Batch and Continuous Cultures under Different H2 Gassing Regimens. Appl Environ Microbiol 73: 740–749. doi: 10.1128/AEM.01885-06
|
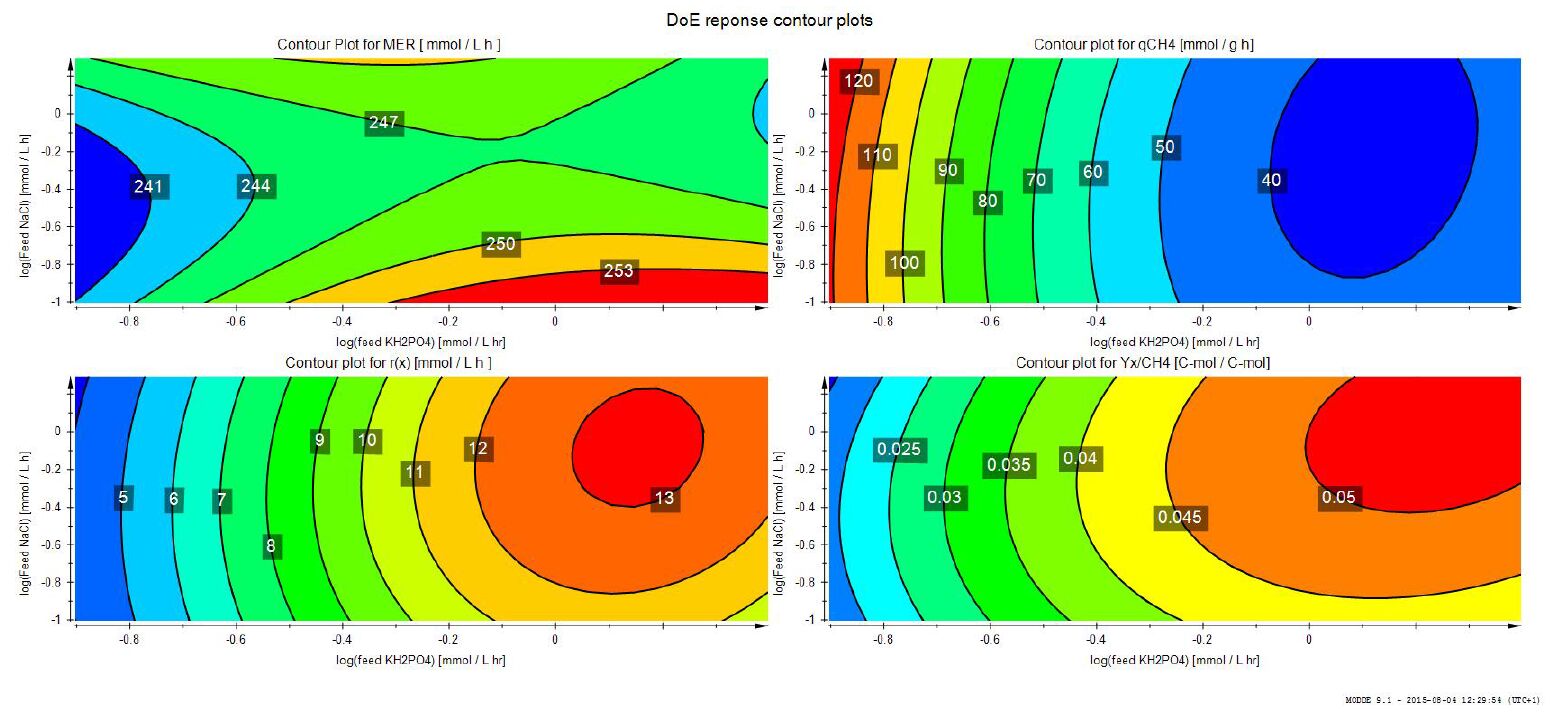
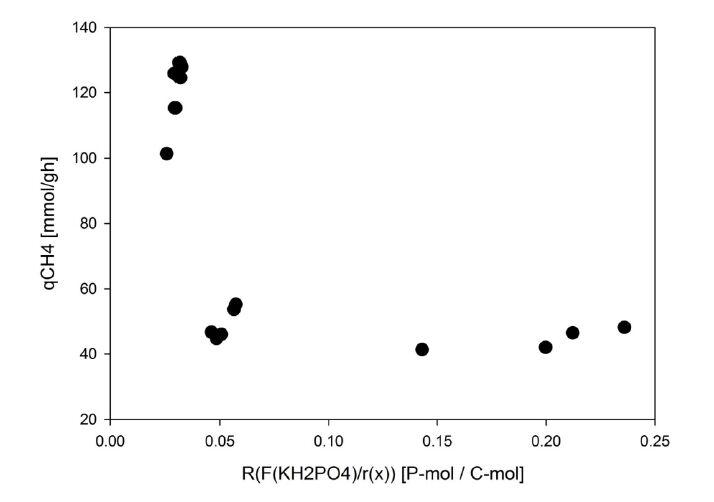
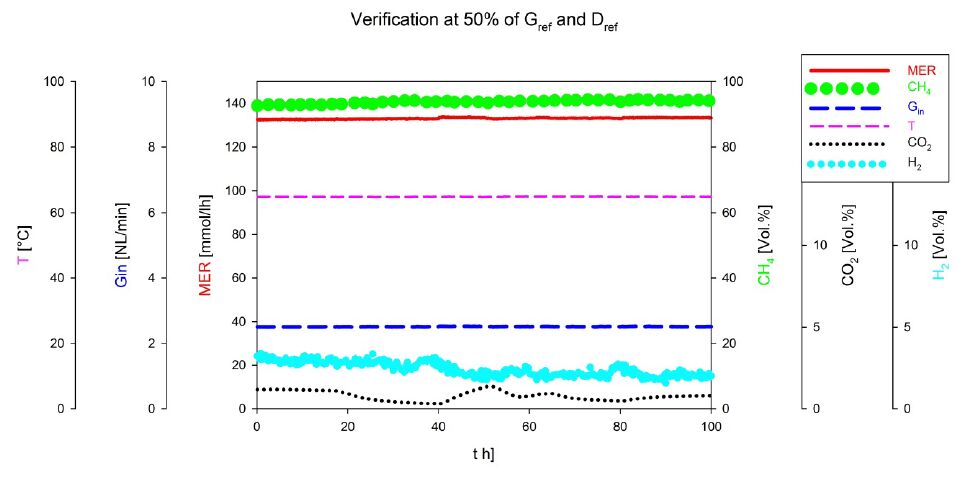
Figures(3) / Tables(2)

Sébastien Bernacchi, Alexander Krajete, Christoph Herwig. Experimental workflow for developing a feed forward strategy to control biomass growth and exploit maximum specific methane productivity of Methanothermobacter marburgensis in a biological methane production process (BMPP)[J]. AIMS Microbiology, 2016, 2(3): 262-277. doi: 10.3934/microbiol.2016.3.262







 DownLoad:
DownLoad: