Citation: Anji Yang, Baojun Song, Sanling Yuan. Noise-induced transitions in a non-smooth SIS epidemic model with media alert[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 745-763. doi: 10.3934/mbe.2021040

| [1] |
M. S. Rahman, M. L. Rahman, Media and education play a tremendous role in mounting aids awareness among married couples in bangladesh, AIDS Res. Ther., 4 (2007), 1–7. doi: 10.1186/1742-6405-4-1

|
| [2] | J. Cui, X. Tao, H. Zhu, An sis infection model incorporating media coverage, Rocky Mt. J. Math., 38 (2008), 1323–1334. |
| [3] |
J. Cui, Y. Sun, H. Zhu, The impact of media on the control of infectious diseases, J. Dyn. Differ. Equ., 20 (2008), 31–53. doi: 10.1007/s10884-007-9075-0
|
| [4] |
L. Wang, D. Zhou, Z. Liu, D. Xu, X. Zhang, Media alert in an sis epidemic model with logistic growth, J. Biol. Dynam., 11 (2017), 120–137. doi: 10.1080/17513758.2016.1181212
|
| [5] |
R. Liu, J. Wu, H. Zhu, Media/psychological impact on multiple outbreaks of emerging infectious diseases, Comput. Math. Methods Med., 8 (2007), 153–164. doi: 10.1080/17486700701425870
|
| [6] |
J. M. Tchuenche, N. Dube, C. P. Bhunu, R. J. Smith, C. T. Bauch, The impact of media coverage on the transmission dynamics of human influenza, BMC Public Health, 11 (2011), 1–16. doi: 10.1186/1471-2458-11-1
|
| [7] |
C. Sun, W. Yang, J. Arino, K. Khan, Effect of media-induced social distancing on disease transmission in a two patch setting, Math. Biosci., 230 (2011), 87–95. doi: 10.1016/j.mbs.2011.01.005
|
| [8] |
A. K. Misra, A. Sharma, J. B. Shukla, Modeling and analysis of effects of awareness programs by media on the spread of infectious diseases, Math. Comput. Model., 53 (2011), 1221–1228. doi: 10.1016/j.mcm.2010.12.005
|
| [9] |
A. Wang, Y. Xiao, A Filippov system describing media effects on the spread of infectious diseases, Nonlinear Anal. Hybr. Syst., 11 (2014), 84–97. doi: 10.1016/j.nahs.2013.06.005
|
| [10] |
Y. Xiao, X. Xu, S. Tang, Sliding mode control of outbreaks of emerging infectious diseases, Bull. Math. Biol., 74 (2012), 2403–2422. doi: 10.1007/s11538-012-9758-5
|
| [11] | R. Gallotti, F. Valle, N. Castaldo, Assessing the risks of "infodemics" in response to COVID-19 epidemics, Nat. Hum. Behav., (2020). |
| [12] |
Y. Zhao, L. Zhang, S. Yuan, The effect of media coverage on threshold dynamics for a stochastic SIS epidemic model, Phys. A, 512 (2018), 248–260. doi: 10.1016/j.physa.2018.08.113
|
| [13] |
A. C. Lowen, J. Steel, Roles of humidity and temperature in shaping in uenza seasonality, J. Virol., 88 (2014), 7692–7695. doi: 10.1128/JVI.03544-13
|
| [14] | I. Bashkirtseva, L. Ryashko, Sensitivity analysis of stochastic attractors and noise-induced transitions for population model with allee effect, Chaos, 21 (2011), 047514. |
| [15] |
I. Bashkirtseva, L. Ryashko, Stochastic sensitivity analysis of noise-induced excitement in a prey–predator plankton system, Front. Life Sci., 5 (2011), 141–148. doi: 10.1080/21553769.2012.702666
|
| [16] | I. Bashkirtseva, L. Ryashko, Stochastic bifurcations and noise-induced chaos in a dynamic prey–predator plankton system, Int. J. Bifurcat. Chaos, 24 (2014), 1450109. |
| [17] | X. Yu, S. Yuan, Asymptotic properties of a stochastic chemostat model with two distributed delays and nonlinear perturbation, Discrete Cont. Dyn. B, 25 (2020), 2373–2390. |
| [18] |
S. Zhao, S. Yuan, H. Wang, Threshold behavior in a stochastic algal growth model with stoichiometric constraints and seasonal variation, J. Differ. Equations, 268 (2020), 5113–5139. doi: 10.1016/j.jde.2019.11.004
|
| [19] |
X. Yu, S. Yuan, T. Zhang, Asymptotic properties of stochastic nutrient-plankton food chain models with nutrient recycling, Nonlinear Anal.-Hybri., 34 (2019), 209–225. doi: 10.1016/j.nahs.2019.06.005
|
| [20] | C. Xu, S. Yuan, T. Zhang, Average break-even concentration in a simple chemostat model with telegraph noise, Nonlinear Anal.-Hybri. 29 (2018), 373–382. |
| [21] |
C. Xu, S. Yuan, Competition in the chemostat: A stochastic multi-species model and its asymptotic behavior, Math. Biosci., 280 (2016), 1–9. doi: 10.1016/j.mbs.2016.07.008
|
| [22] | C. Xu, S. Yuan, T. Zhang, Competitive exclusion in a general multi-species chemostat model with stochastic perturbations, Bull. Math. Biol., DOI: 10.1007/s11538-020-00843-7. |
| [23] |
Y. Zhao, S. Yuan, J. Ma, Survival and stationary distribution analysis of a stochastic competitive model of three species in a polluted environment, Bull. Math. Biol., 77 (2015), 1285–1326. doi: 10.1007/s11538-015-0086-4
|
| [24] |
D. Wu, H. Wang, S. Yuan, Stochastic sensitivity analysis of noise-induced transitions in a predator-prey model with environmental toxins, Math. Biosci. Eng., 16 (2019), 2141–2153. doi: 10.3934/mbe.2019104
|
| [25] | C. Xu, S. Yuan, T. Zhang, Stochastic sensitivity analysis for a competitive turbidostat model with inhibitory nutrients, Int. J. Bifurcat. Chaos, 26 (2016), 707–723. |
| [26] | L. Gammaitoni, P. Jung, F. Marchesoni, Stochastic resonance, Rev. Mod. Phys., 70 (1998), 223. |
| [27] | W. Horsthemke, Noise induced transitions, Non-Equi. Dyna. Chem. Sys., (1984), 150–160. |
| [28] | K. Matsumoto, I. Tsuda, Noise-induced order, J. Stat. Phys., 31 (1983), 87–106. |
| [29] | J. Gao, S. Hwang, J. Liu, When can noise induce chaos? Phys. Rev. Lett., 82 (1999), 1132–1135. |
| [30] | M. A. Zaks, X. Sailer, L. S. Geier, A. Neiman, Noise induced complexity: From subthreshold oscillations to spiking in coupled excitable systems, Chaos, 15 (2005), 026117. |
| [31] | S. Kim, S. H. Park, C. S. Ryu, Colored-noise-induced multistability in nonequilibrium phase transitions, Phys. Rev. E, 58 (1998), 7994-7997. |
| [32] |
S. L. Souza, A. M. Batista, I. L. Caldas, R. L. Viana, T. Kapitaniak, Noise-induced basin hopping in a vibro-impact system, Chaos Soliton. Fract., 32 (2007), 758–767. doi: 10.1016/j.chaos.2005.11.056
|
| [33] |
M. I. Dykman, R. Mannella, P. V. E. McClintock, N. Stocks, Fluctuation-induced transitions between periodic attractors: Observation of supernarrow spectral peaks near a kinetic phase transition, Phys. Rev. Lett., 65 (1990), 48–51. doi: 10.1103/PhysRevLett.65.48
|
| [34] | S. Yuan, D. Wu, G. Lan, H. Wang, Noise-induced transitions in a nonsmooth producer-grazer model with stoichiometric constraints, Bull. Math. Biol., 82 (2020), 55. |
| [35] | S. Kraut, U. Feudel, Multistability, noise, and attractor hopping: The crucial role of chaotic saddles, Phys. Rev. E, 66 (2002), 015207. |
| [36] | I. Bashkirtseva, L. Ryashko, I.Tsvetkov, Sensitivity analysis of stochastic equilibria and cycles for the discrete dynamic systems, Dyna. Cont. Dis. Imp. Syst., 17 (2010), 501–515. |
| [37] | I. Bashkirtseva, T. Ryazanova, L. Ryashko, Confidence domains in the analysis of noiseinduced transition to chaos for Goodwin model of business cycles, Int. J. Bifurc. Chaos, 24 (2014), 1440020. |


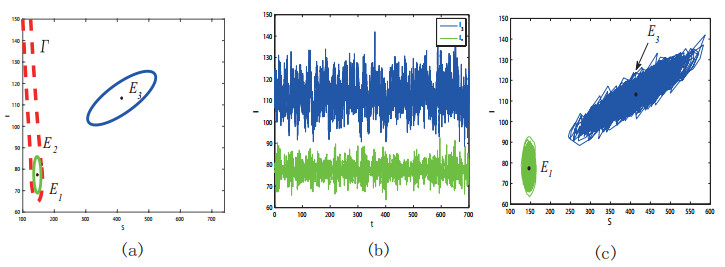
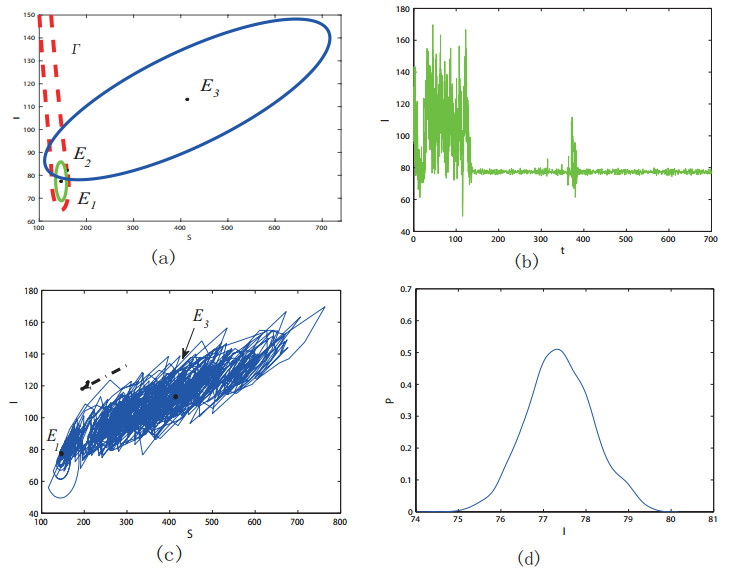
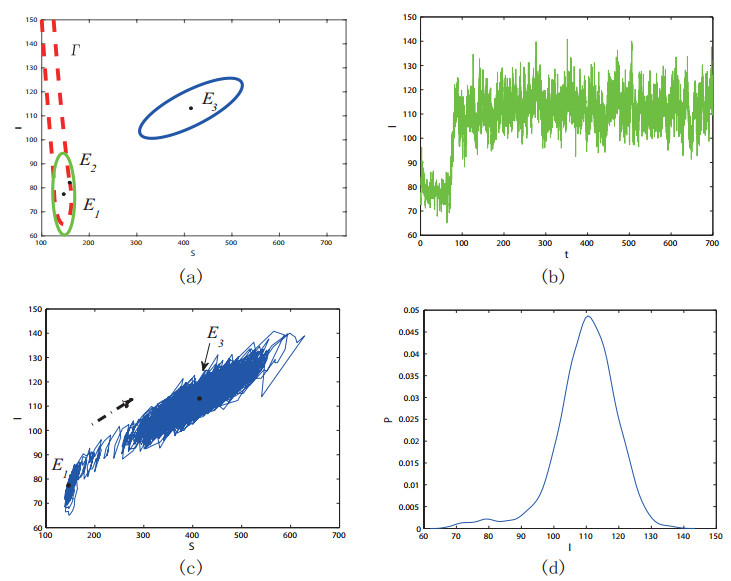
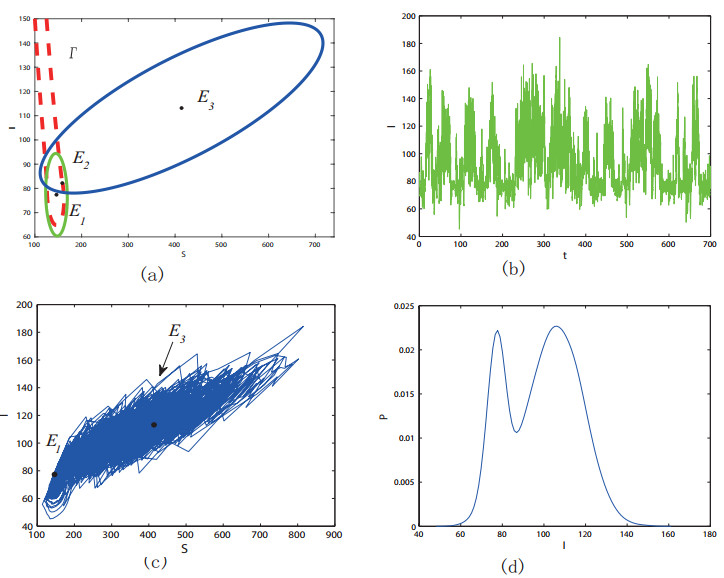
Figures(11) / Tables(1)

Anji Yang, Baojun Song, Sanling Yuan. Noise-induced transitions in a non-smooth SIS epidemic model with media alert[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 745-763. doi: 10.3934/mbe.2021040







 DownLoad:
DownLoad: