Citation: Raimund Bürger, Gerardo Chowell, Elvis Gavilán, Pep Mulet, Luis M. Villada. Numerical solution of a spatio-temporal predator-prey model with infected prey[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 438-473. doi: 10.3934/mbe.2019021

| [1] | L.J.S. Allen, B.M. Bolker, Y. Lou, and A.L. Nevai, Asymptotic profiles of the steady states for an SIS epidemic patch model, SIAM J. Appl. Math., 67 (2007), 1283–1309. |
| [2] | J. Arino, Diseases in metapopulations. In Z. Ma, Y. Zhou, and J. Wu (Eds.), Modeling and Dynamics of Infectious Diseases, Higher Education Press, Beijing, 2009, 64–122. |
| [3] | J. Arino, J.R. Davis, D. Hartley, R. Jordan, J.M. Miller, and P. van den Driessche, A multi-species epidemic model with spatial dynamics, Math. Med. Biol., 22 (2005), 129–142. |
| [4] | U. Ascher, S. Ruuth, and J. Spiteri, Implicit-explicit Runge-Kutta methods for time dependent partial differential equations, Appl. Numer. Math., 25 (1997), 151–167. |
| [5] | S. Boscarino, R. Bürger, P. Mulet, G. Russo, and L.M. Villada, Linearly implicit IMEX Runge- Kutta methods for a class of degenerate convection-diffusion problems, SIAM J. Sci. Comput., 37 (2015), B305–B331. |
| [6] | S. Boscarino, F. Filbet, and G. Russo, High order semi-implicit schemes for time dependent partial differential equations, J. Sci. Comput., 68 (2016), 975–1001. |
| [7] | S. Boscarino, P.G. LeFloch, and G. Russo, High-order asymptotic-preserving methods for fully nonlinear relaxation problems, SIAM J. Sci. Comput., 36 (2014), A377–A395. |
| [8] | S. Boscarino and G. Russo, On a class of uniformly accurate IMEX Runge-Kutta schemes and applications to hyperbolic systems with relaxation, SIAM J. Sci. Comput., 31 (2009), 1926–1945. |
| [9] | S. Boscarino and G. Russo, Flux-explicit IMEX Runge-Kutta schemes for hyperbolic to parabolic relaxation problems, SIAM J. Numer. Anal., 51 (2013), 163–190. |
| [10] | F. Brauer and C. Kribs, Dynamical Systems for Biological Modeling: An Introduction, CRC Press, Boca Raton, FL, USA, 2016. |
| [11] | R. Bürger, G. Chowell, E. Gavilán, P. Mulet, and L.M. Villada, Numerical solution of a spatiotemporal gender-structured model for hantavirus infection in rodents, Math. Biosci. Eng., 15 (2018), 95–123. |
| [12] | R. Bürger, G. Chowell, P. Mulet, and L.M. Villada, Modelling the spatial-temporal progression of the 2009 A/H1N1 influenza pandemic in Chile, Math. Biosci. Eng., 13 (2016), 43–65. |
| [13] | J. Chattopadhyay and O. Arino, A predator-prey model with disease in the prey, Nonlin. Anal., 36 (1999), 747–766. |
| [14] | J. Chattopadhyay and N. Bairagi, Pelicans at risk in Salton sea-an eco-epidemiological model, Ecol. Model., 136 (2001), 103–112. |
| [15] | J. Chattopadhyay, P.D.N. Srinivasu, and N. Bairagi, Pelicans at risk in Salton sea-an ecoepidemiological model-II, Ecol. Model., 167 (2003), 199-211. |
| [16] | R.M. Colombo and E. Rossi, Hyperbolic predators versus parabolic preys, Commun. Math. Sci., 13 (2015), 369–400. |
| [17] | M. Crouzeix, Une méthode multipas implicite-explicite pour l'approximation des équations d'évolution paraboliques, Numer. Math., 35 (1980), 257–276. |
| [18] | O. Diekmann, H. Heesterbeek, and T. Britton, Mathematical Tools for Understanding Infectious Disease Dynamics, Princeton Series in Theoretical and Computational Biology, Princeton University Press, 2012. |
| [19] | R. Donat and I. Higueras, On stability issues for IMEX schemes applied to 1D scalar hyperbolic equations with stiff reaction terms, Math. Comp., 80 (2011), 2097–2126. |
| [20] | L. Edelstein-Keshet, Mathematical Models in Biology, reprint, SIAM, 2005. |
| [21] | I.M. Foppa, A Historical Introduction to Mathematical Modeling of Infectious Diseases, Academic Press, London, UK, 2016. |
| [22] | D. Greenhalgh and M. Haque, A predator-prey model with disease in the prey species only, Math. Meth. Appl. Sci., 30 (2007), 911–929. |
| [23] | D. Greenhalgh, Q.J.A. Khan, and J.S. Pettigrew, An eco-epidemiological predator-prey modelmwhere predators distinguish between susceptible and infected prey, Math. Meth. Appl. Sci., 40 (2017), 146–166. |
| [24] | K.P. Hadeler and H.I. Freedman, Predator-prey populations with parasitic infection, J. Math. Biol., 27 (1989), 609–631. |
| [25] | H.W. Hethcote, W. Wang, L. Han, and Z. Ma, A predator-prey model with infected prey, Theor. Population Biol., 66 (2004), 259–268. |
| [26] | G.S. Jiang and C.W. Shu, Efficient implementation of weighted ENO schemes, J. Comput. Phys., 126 (1996), 202–228. |
| [27] | Y. Katznelson, An Introduction to Harmonic Analysis, Third Ed., Cambridge University Press, Cambridge, UK, 2004. |
| [28] | C.A. Kennedy and M.H. Carpenter, Additive Runge-Kutta schemes for convection-diffusionreaction equations, Appl. Numer. Math., 44 (2003), 139–181. |
| [29] | Y. Kuang and E. Beretta, Global qualitative analysis of a ratio-dependent predator-prey system, J. Math. Biol., 36 (1998), 389–406. |
| [30] | K. Kundu and J. Chattopadhyay, A ratio-dependent eco-epidemiological model of the Salton Sea, Math. Meth. Appl. Sci., 29 (2006), 191–207. |
| [31] | P.H. Leslie and J.C. Gower, The properties of a stochastic model for the predator-prey type of interaction between two species, Biometrika, 47 (1960), 219–234. |
| [32] | W.M. Liu, H.W. Hethcote, and S.A. Levin, Dynamical behavior of epidemiological models with nonlinear incidence rates, J. Math. Biol., 25 (1987), 359–380. |
| [33] | W.M. Liu, S.A. Levin, and Y. Iwasa, Influence of nonlinear incidence rates upon the behavior of SIRS epidemiological models, J. Math. Biol., 23 (1986), 187–204. |
| [34] | X.D. Liu, S. Osher, and T. Chan, Weighted essentially non-oscillatory schemes, J. Comput. Phys., 115 (1994), 200–212. |
| [35] | H. Malchow, S.V. Petrovskii, and E. Venturino, Spatial Patterns in Ecology and Epidemiology: Theory, Models, and Simulation. Chapman & Hall/CRC, Boca Raton, FL, USA, 2008. |
| [36] | J.D. Murray, Mathematical Biology II: Spatial Models and Biomedical Applications. Third Edition. Springer, New York, 2003. |
| [37] | A. Okubo and S.A. Levin, Diffusion and Ecological Problems: Modern Perspectives. Second Edition, Springer-Verlag, New York, 2001. |
| [38] | L. Pareschi and G. Russo, Implicit-Explicit Runge-Kutta schemes and applications to hyperbolic systems with relaxation, J. Sci. Comput., 25 (2005), 129–155. |
| [39] | E. Rossi and V. Schleper, Convergence of a numerical scheme for a mixed hyperbolic-parabolic system in two space dimensions, ESAIM Math. Modelling Numer. Anal., 50 (2016), 475–497. |
| [40] | R. Sarkar, J. Chattopadhyay, and N. Bairagi, Effects of environmental fluctuation on an ecoepidemiological model of the Salton Sea, Environmetrics, 12 (2001), 289–300. |
| [41] | L. Sattenspiel, The Geographic Spread of Infectious Diseases: Models and Applications, Princeton Series in Theoretical and Computational Biology, Princeton University Press, 2009. |
| [42] | C.W. Shu and S. Osher, Efficient implementation of essentially non-oscillatory shock-capturing schemes, II, J. Comput. Phys., 83 (1988), 32–78. |
| [43] | S.W. Smith, Digital Signal Processing: A Practical Guide for Engineers and Scientists. Demystifying technology series: by engineers, for engineers. Newnes, 2003. |
| [44] | S.H. Strogatz, Nonlinear Dynamics and Chaos: with Applications to Physics, Biology, Chemistry, and Engineering, Second Edition, CRC Press, Boca Raton, FL, 2015. |
| [45] | G. Q. Sun, Pattern formation of an epidemic model with diffusion, Nonlinear Dynamics, 69 (2012), 1097–1104. |
| [46] | P. van den Driessche, Deterministic compartmental models: extensions of basic models. In F. Brauer, P. van den Driessche, and J. Wu (Eds.), Mathematical Epidemiology, Springer-Verlag, Berlin, 2008, 147–157. |
| [47] | P. van den Driessche, Spatial structure: patch models. In F. Brauer, P. van den Driessche and J.Wu (Eds.), Mathematical Epidemiology, Springer-Verlag, Berlin, 2008, 179–189. |
| [48] | E. Vynnycky and R.E. White, An Introduction to Infectious Disease Modelling, Oxford University Press, 2010. |
| [49] | J. Wu, Spatial structure: partial differential equations models. In F. Brauer, P. van den Driessche and J. Wu (Eds.), Mathematical Epidemiology, Springer-Verlag, Berlin, 2008, 191–203. |
| [50] | P. Yu, Closed-form conditions of bifurcation points for general differential equations, Internat. J. Bifur. Chaos Appl. Sci. Engrg., 15 (2005), 1467–1483. |
| [51] | Y.T. Zhang and C.W. Shu, ENO and WENO schemes. Chapter 5 in R. Abgrall and C.W. Shu (eds.), Handbook of Numerical Methods for Hyperbolic Problems Basic and Fundamental Issues. Handbook of Numerical Analysis vol. 17, North Holland (2016), 103–122. |
| [52] | X. Zhong, Additive semi-implicit Runge-Kutta methods for computing high-speed nonequilibrium reactive flows, J. Comput. Phys., 128 (1996), 19–31. |
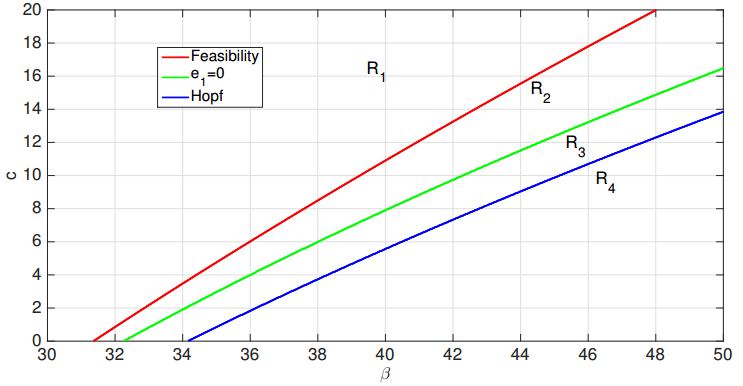
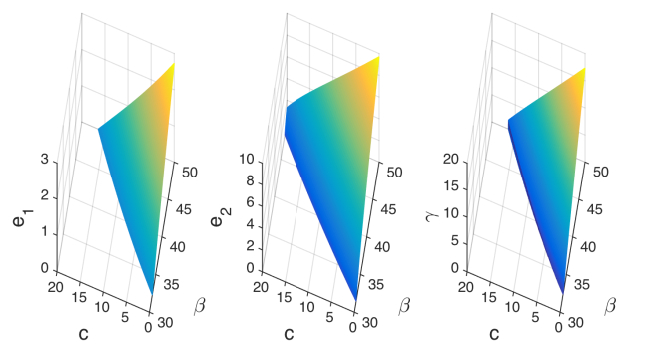
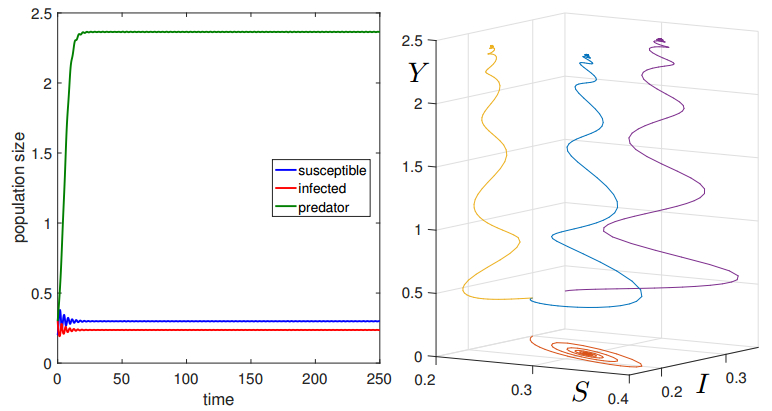
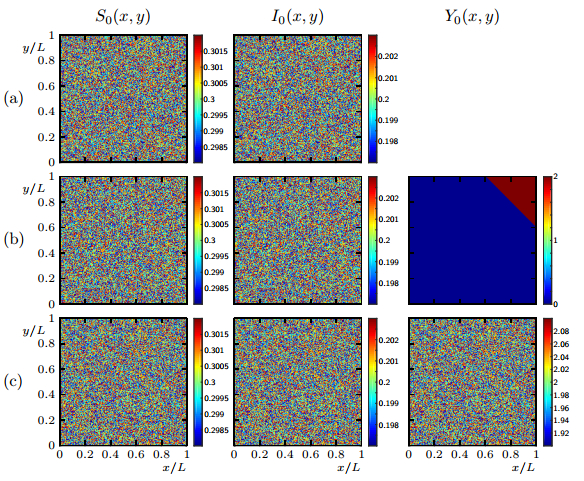
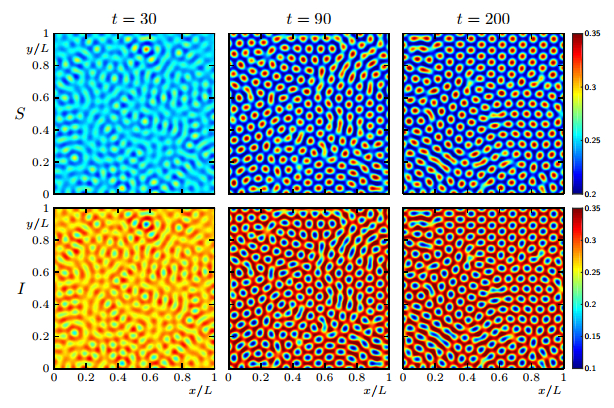
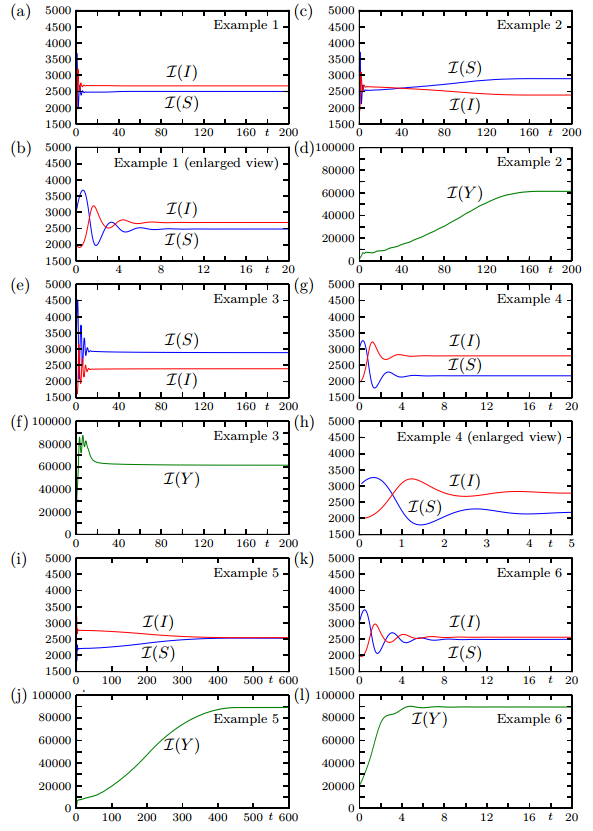
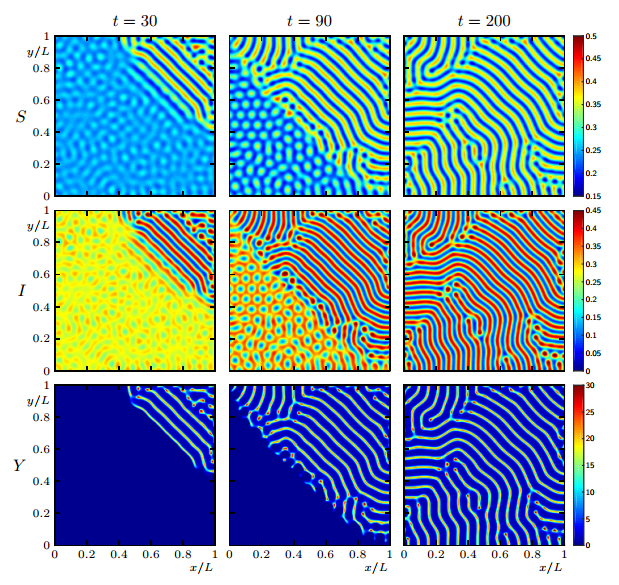
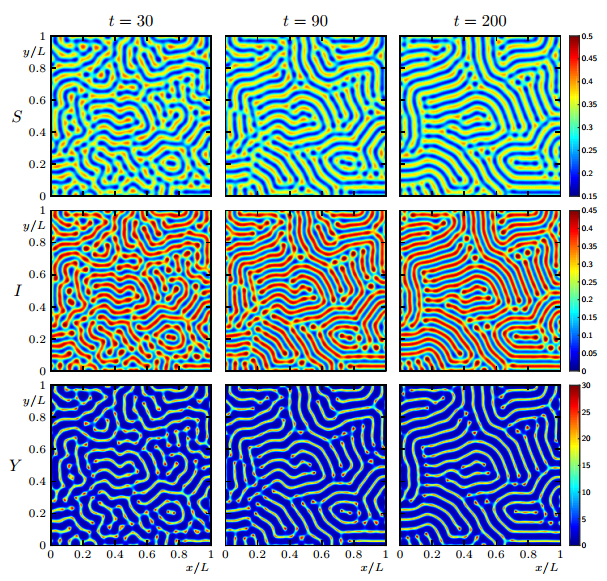
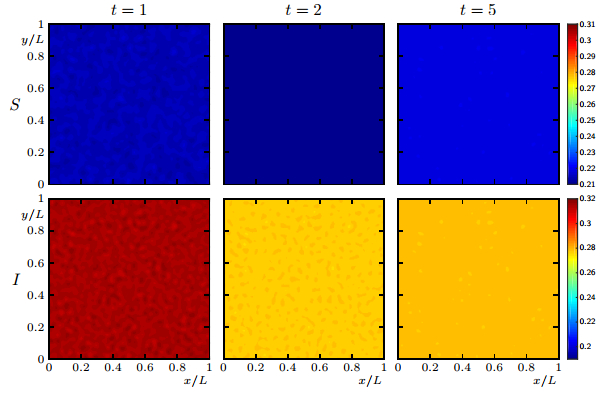
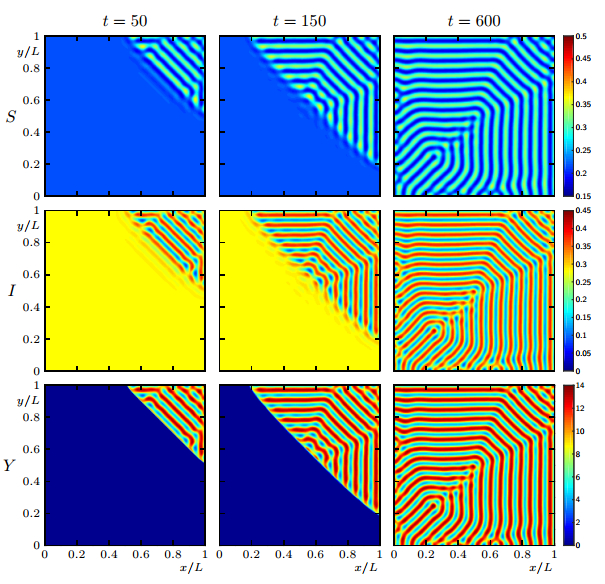
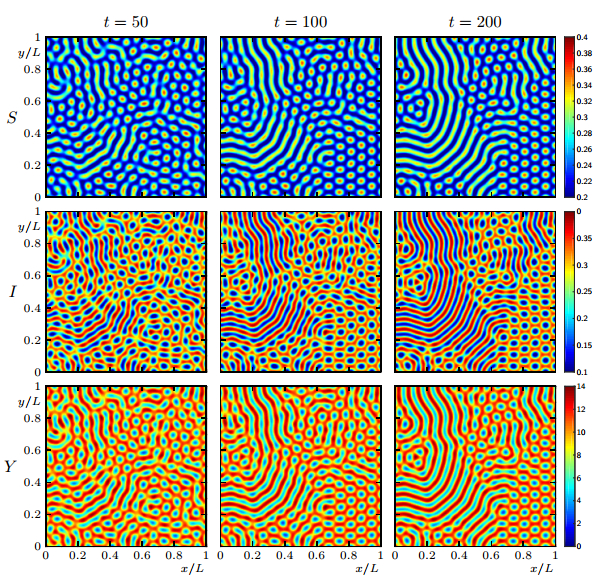
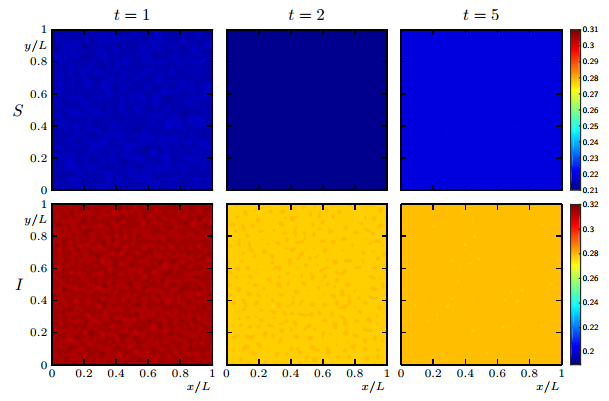
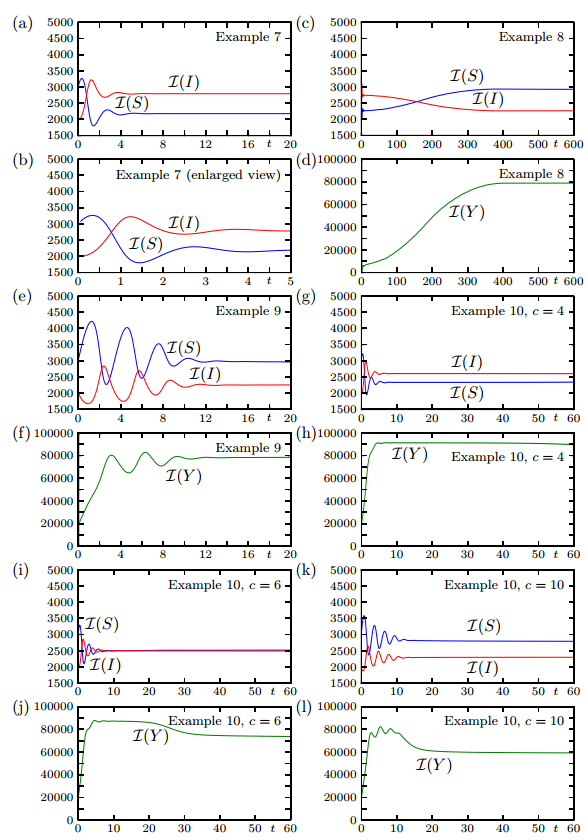
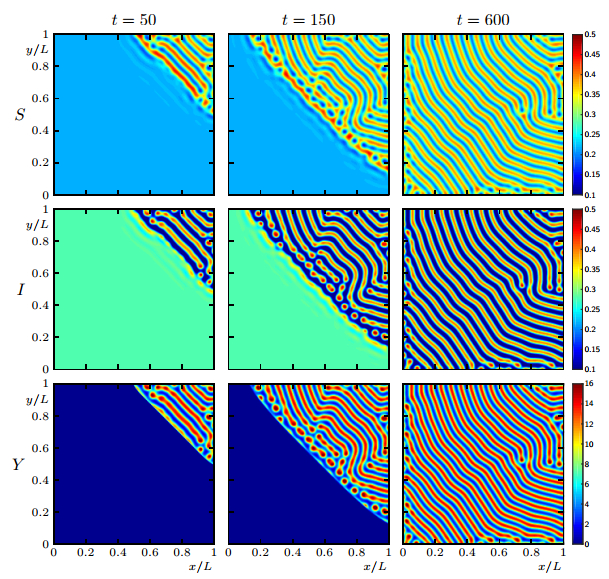
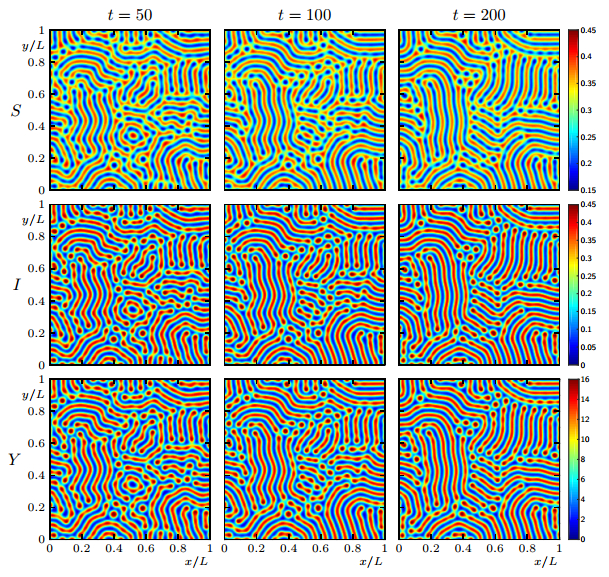
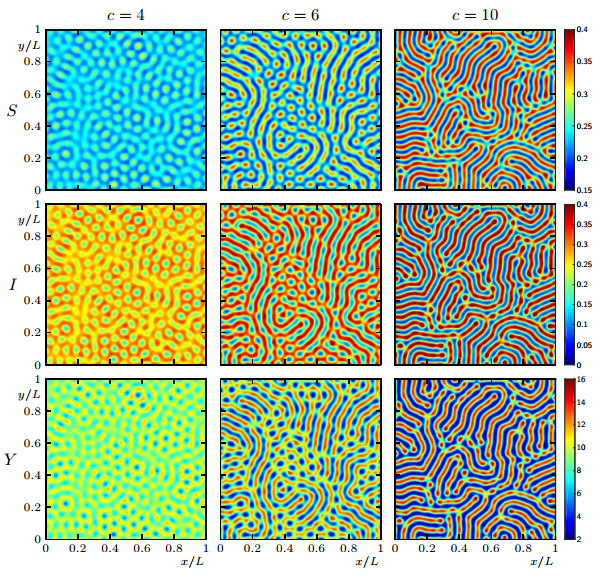
Figures(19)

Raimund Bürger, Gerardo Chowell, Elvis Gavilán, Pep Mulet, Luis M. Villada. Numerical solution of a spatio-temporal predator-prey model with infected prey[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 438-473. doi: 10.3934/mbe.2019021







 DownLoad:
DownLoad: