Citation: Konstantin E. Starkov, Giovana Andres Garfias. Dynamics of the tumor-immune-virus interactions: Convergence conditions to tumor-only or tumor-free equilibrium points[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 421-437. doi: 10.3934/mbe.2019020

| [1] | Ž. Bajzer, T. Carr, K. Josić, S.J Russell and D. Dingli, Modeling of cancer virotherapy with recombinant measles viruses, J. Theor. Biol. 252 (2008), 109–122. |
| [2] | M. Biesecker, J.H. Kimn, H. Lu, D. Dingli and Ž. Bajzer, Optimization of virotherapy for cancer, Bulletin Math. Biol., 72 (2010), 469–489. |
| [3] | B.W. Bridle, K.B. Stephenson, J.E. Boudreau, S. Koshy, N. Kazdhan, E. Pullenayegum, J. Brunellière, J.L. Bramson, B.D. Lichty and Y. Wan, Potentiating cancer immunotherapy using an oncolytic virus, Mol. Ther., 18 (2010), 1430–1439. |
| [4] | R.M. Diaz, F. Galivo, T. Kottke, P.Wongthida, J. Qiao, J. Thompson, M. Valdes, G. Barber and R.G Vile, Oncolytic immunovirotherapy for melanoma using vesicular stomatitis virus, Cancer Res., 67 (2007), 2840–2848. |
| [5] | R. Eftimie, J. Dushoff, B.M. Bridle, J.L. Bramson and D.J.D. Earn, Multi-stability and multiinstability phenomena in a mathematical model of tumor-immune-virus interactions, Bulletin Math. Biol., 73 (2011), 2932–2961. |
| [6] | R. Eftimie, C.K. Macnamara, J. Dushoff, J.L. Bramson and D.J.D. Earn, Bifurcations and chaotic dynamics in a tumour-immune-virus system, Math. Modell. Nat. Pheno., 11 (2016), 65–85. |
| [7] | M.W. Hirsch, Systems of differential equations that are competitive or cooperative II: Convergence almost everywhere, SIAM J. Math. Analysis, 16 (1985), 423–439. |
| [8] | B.Y. Hwang and D.V. Schaffer, Engineering a serum-resistant and thermostable vesicular stomatitis virus G glycoprotein for pseudotyping retroviral and lentiviral vectors, Gene Ther., 20 (2013), 807– 815. |
| [9] | A.P. Krishchenko, Estimation of domains with cycles, Computer Math. Appl., 34 (1997), 325–332. |
| [10] | A.P. Krishchenko, Localization of invariant compact sets of dynamical systems, Differ. Equations, 41 (2005), 1669–1676. |
| [11] | A.P. Krishchenko and K.E. Starkov, Localization of compact invariant sets of the Lorenz system, Phys. Lett. A, 353 (2006), 383–388. |
| [12] | A.P. Krishchenko and K.E. Starkov, On the global dynamics of a chronic myelogenous leukemia model, Commun. Nonlin. Sci. Numer. Simul., 33 (2016), 174–183. |
| [13] | A.P. Krishchenko and K.E. Starkov, The four-dimensional Kirschner-Panetta type cancer model: how to obtain tumor eradication? Math. Biosci. Eng., 15 (2018), 1243–1254. |
| [14] | S. Varghese and S.D. Rabkin, Oncolytic herpes simplex virus vectors for cancer virotherapy, Cancer Gene Ther., 9 (2002), 967–978. |
| [15] | M.Y. Li and S.J. Muldowney, On R.A. Smith's autonomous convergence theorem, Rocky Mountain J. Math., 25 (1995), 365–379. |
| [16] | R.A. Smith, Some applications of Hausdorff dimension inequalities for ordinary differential equations, Proceed. Royal Soc. Edinburgh Sec. A: Math., 104 (1986), 235–259. |
| [17] | K.E. Starkov, On dynamic tumor eradication conditions under combined chemical/anti-angiogenic therapies, Phys. Lett. A, 382 (2018), 387–393. |
| [18] | K.E. Starkov and S. Bunimovich-Mendrazitsky, Dynamical properties and tumor clearance conditions for a nine-dimensional model of bladder cancer immunotherapy, Math. Biosci. Eng., 13 (2016), 1059–1075. |
| [19] | K.E. Starkov and L. Jimenez Beristain, Dynamic analysis of the melanoma model: from cancer persistence to its eradication, Internat. J. Bifur. Chaos Appl. Sci. Engrg., 27 (2017), 1750151-1– 1750151-11. |
| [20] | K.E. Starkov and A.P. Krishchenko, Ultimate dynamics of the Kirschner-Panetta model: Tumor eradication and related problems, Phys. Lett. A, 381 (2017), 3409–3016. |
| [21] | H.R. Thieme, Asymptotically autonomous differential equations in the plane, Rocky Mountain J. Math., 34 (1994), 351–380. |
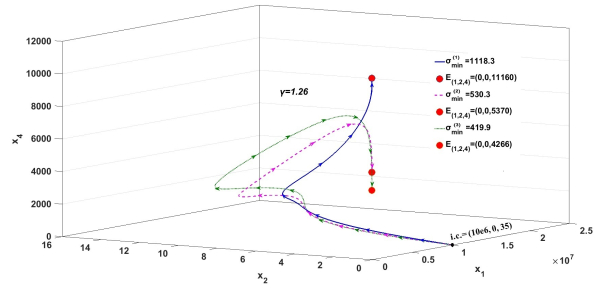
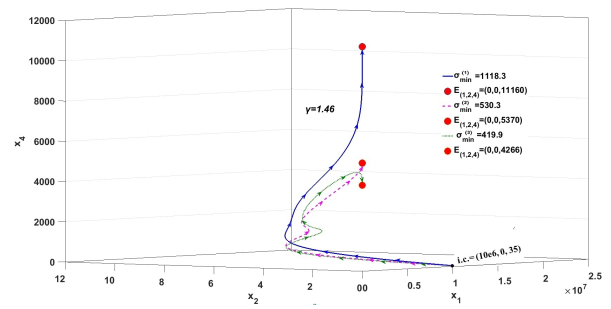
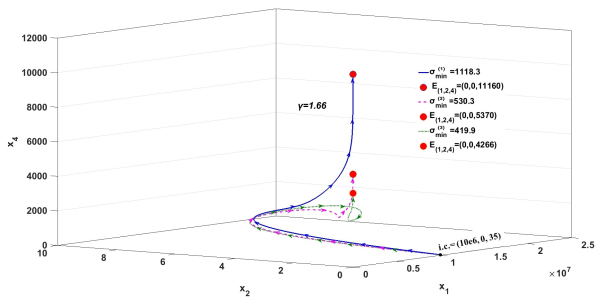
Figures(3)

Konstantin E. Starkov, Giovana Andres Garfias. Dynamics of the tumor-immune-virus interactions: Convergence conditions to tumor-only or tumor-free equilibrium points[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 421-437. doi: 10.3934/mbe.2019020







 DownLoad:
DownLoad: