In this paper, we establish a non-autonomous Hassell-Varley-Holling type predator-prey system with mutual interference. We construct some sufficient conditions for the permanence, extinction and globally asymptotic stability of system by use of the comparison theorem and an appropriate Liapunov function. Then the sufficient and necessary conditions for a periodic solution of the system are obtained via coincidence degree theorem. Finally, the correctness of the previous conclusions are demonstrated by some numerical cases.
Citation: Luoyi Wu, Hang Zheng, Songchuan Zhang. Dynamics of a non-autonomous predator-prey system with Hassell-Varley-Holling Ⅱ function response and mutual interference[J]. AIMS Mathematics, 2021, 6(6): 6033-6049. doi: 10.3934/math.2021355
In this paper, we establish a non-autonomous Hassell-Varley-Holling type predator-prey system with mutual interference. We construct some sufficient conditions for the permanence, extinction and globally asymptotic stability of system by use of the comparison theorem and an appropriate Liapunov function. Then the sufficient and necessary conditions for a periodic solution of the system are obtained via coincidence degree theorem. Finally, the correctness of the previous conclusions are demonstrated by some numerical cases.

| [1] | C. Holling, The functional response of predator to prey density and its role in mimicy and population regulation, J. Men. Ent. Sec. Can., 45 (1965), 1–60. |
| [2] |
F. Wei, Existence of multiple positive periodic solutions to a periodic predator-prey system with harvesting terms and Holling Ⅲ type functional response, Commun. Nonlinear Sci. Numer. Simul., 16 (2011), 2130–2138. doi: 10.1016/j.cnsns.2010.08.028

|
| [3] |
F. Chen, Y. Chen, J. Shi, Stability of the boundary solution of a nonautonomous predator-prey system with the Beddington-DeAngelis functional response, J. Math. Anal. Appl., 344 (2008), 1057–1067. doi: 10.1016/j.jmaa.2008.03.050
|
| [4] |
H. Alsakaji, S. Kundu, F. Rihan, Delay differential model of one-predator two-prey system with Monod-Haldane and holling type Ⅱ functional responses, Appl. Math. Comput., 397 (2021), 125919. doi: 10.1016/j.amc.2020.125919
|
| [5] |
Z. Du, Z. Feng, Periodic solutions of a neutral impulsive predator-prey model with Beddington-DeAngelis functional response with delays, J. Comput. Appl. Math., 258 (2014), 87–98. doi: 10.1016/j.cam.2013.09.008
|
| [6] |
H. Guo, X. Chen, Existence and global attractivity of positive periodic solution for a Volterra model with mutual interference and Beddington-DeAngelis functional response, Appl. Math. Comput., 217 (2011), 5830–5837. doi: 10.1016/j.amc.2010.12.065
|
| [7] | J. Song, Y. Xia, Y. Bai, Y. Cai, D. O'Regan, A non-autonomous Leslie-Gower model with Holling type Ⅳ functional response and harvesting complexity, Adv. Differ. Equations, 1 (2019), 1–12. |
| [8] |
F. Wei, Q. Fu, Hopf bifurcation and stability for predator-prey systems with Beddington-DeAngelis type functional response and stage structure for prey incorporating refuge, Appl. Math. Model, 40 (2016), 126–134. doi: 10.1016/j.apm.2015.04.042
|
| [9] |
S. Yuan, Y. Song, Stability and Hopf bifurcations in a delayed Leslie-Gower predator-prey system, J. Math. Anal. Appl., 355 (2009), 82–100. doi: 10.1016/j.jmaa.2009.01.052
|
| [10] | F. Rihan, H. Alsakaji, C. Rajivganthi, Stability and hopf bifurcation of three-species prey-predator System with time delays and Allee Effect, Complexity, (2020), 7306412. |
| [11] |
H. Qi, X. Meng, Threshold behavior of a stochastic predator-prey system with prey refuge and fear effect, Appl. Math. Lett., 113 (2021), 106846. doi: 10.1016/j.aml.2020.106846
|
| [12] |
W. Lu, Y. Xia, Y. Bai, Periodic solution of a stage-structured predator-prey model incorporating prey refuge, Math. Biosci. Eng., 17 (2020), 3160–3174. doi: 10.3934/mbe.2020179
|
| [13] |
Y. Xia, J. Cao, S. Cheng, Multiple periodic solutious of prelayed stage-tructured predator-prey model with nonmonotone functional responses, Appl. Math. Model., 31 (2007), 1947–1959. doi: 10.1016/j.apm.2006.08.012
|
| [14] |
Z. Li, K. Zhao, Y. Li, Multiple positive periodic solutions for a non-autonomous stage-structured predator-prey system with harvesting terms, Commun. Nonlinear Sci. Numer. Simul., 15 (2010), 2140–2148. doi: 10.1016/j.cnsns.2009.08.019
|
| [15] |
Y. Chen, Multiple periodic solution of delayed predator-prey systems with type Ⅳ functional responses, Nonlinear Anal., 5 (2004), 45–53. doi: 10.1016/S1468-1218(03)00014-2
|
| [16] |
S. Yuan, F. Zhang, Stability and global Hopf bifurcation in a delayed predator-prey system, Nonlinear Anal. Real World Appl., 11 (2010), 959–977. doi: 10.1016/j.nonrwa.2009.01.038
|
| [17] |
L. Nie, Z. Teng, L. Hu, J. Peng, The dynamics of a Lotka-Volterra predator-prey model with state dependent impulsive harvest for predator, Biosystems, 98 (2009), 67–72. doi: 10.1016/j.biosystems.2009.06.001
|
| [18] | F. Chen, X. Chen, Y. Zhang, Positive periodic solution of a delayed predator-prey system with Holling type Ⅱ functional response and stage structure for predator, Acta Math. Sci., 26 (2006), 93–103. |
| [19] |
Y. Song, X. Tang, Stability, Steady-State Bifurcations and Turing Patterns in a Predator-Prey Model with Herd Behavior and Prey-taxis, Stud. Appl. Math., 139 (2017), 371–404. doi: 10.1111/sapm.12165
|
| [20] | T. Ma, X. Meng, Z. Chang, Dynamics and optimal harvesting control for a stochastic one-predator-two-prey time delay system with jumps, Complexity, 2019 (2019), 5342031. |
| [21] |
G. Liu, H. Qi, Z. Chang, X. Meng, Asymptotic stability of a stochastic may mutualism system, Comput. Math. Appl., 79 (2020), 735–745. doi: 10.1016/j.camwa.2019.07.022
|
| [22] |
M. Hassell, Density dependence in single-species population, J. Anim. Ecol., 44 (1975), 283–295. doi: 10.2307/3863
|
| [23] |
X. Chen, Z. Du, Existence of positive periodic solutions for a neutral delay predator-prey model with Hassell-Varley type functional response and impulse, Theory Dyn. Syst., 17 (2018), 67–80. doi: 10.1007/s12346-017-0223-6
|
| [24] | H. Liu, F. Meng, X. He, Existence of positive periodic solutions for a predator-prey system of Holling type Ⅳ function response with mutual interference and impulsive effects, Discrete Dyn. Nat. Soc., 2015 (2015), 1–12. |
| [25] | J. Fu, L. Chen, Qualitative analysis of a class predator-prey model with mutual interference, J. Systems Sci. Math. Sci., 37 (2017), 1166–1178. |
| [26] |
M. Hassell, G. Varley, New inductive population model for insect parasites and its bearing on biological control, Nature, 223 (1969), 1133–1137. doi: 10.1038/2231133a0
|
| [27] |
R. Arditi, H. Ackakaya, Underestimation of mutual interference of predators, Oeclogia., 83 (1990), 358–361. doi: 10.1007/BF00317560
|
| [28] |
W. Satherland, Aggregation and the idea free distribution, J. Anim. Ecol., 52 (1983), 821–828. doi: 10.2307/4456
|
| [29] | S. Hsu, T. Hwang, Y. Kuang, Global dynamics of a predator-prey model with Hassell-Varley type functional response, Discrete Contin. Dyn. Syst., 10 (2008), 857–871. |
| [30] | R. Wu, L. Li, M. DelaSen, Permanence and Global Attractivity of the Discrete Predator-Prey System with Hassell-Varley-Holling Ⅲ Type Functional Response, Discrete Dyn. Nat. Soc., 2013 (2013), 295–321. |
| [31] | M. Zhong, X. Liu, Dynamical analysis of a predator-prey system with Hassell-Varley-Holling function response, J. Math. Phys., 31 (2011), 1295–1310. |
| [32] |
M. Fan, Q. Wang, X. Zou, Dynamics of a non-autonomous ratio-dependent predator-prey system, Proc. R. Soc. Edinburgh Sect. A: Math., 133 (2003), 97–118. doi: 10.1017/S0308210500002304
|
| [33] | M. Fan, K. Wang, Global existence of positive periodic solution of a Predator-Prey system with Holling Ⅱ function response, Acta Math. Sci. Ser. A Chin. Ed., 21 (2011), 492–497. |
| [34] |
S. Zhang, X. Meng, T. Zhang, Dynamics analysis and numerical simulations of a stochastic non-autonomous predator-prey system with impulsive effects, Nonlinear Anal. Hybri., 26 (2017), 19–37. doi: 10.1016/j.nahs.2017.04.003
|
| [35] | R. Gaines, T. Mawhin, Coincidence Degree and Nonlinear Differential Equations, Berlin: Springer-Verlag, 1977. |
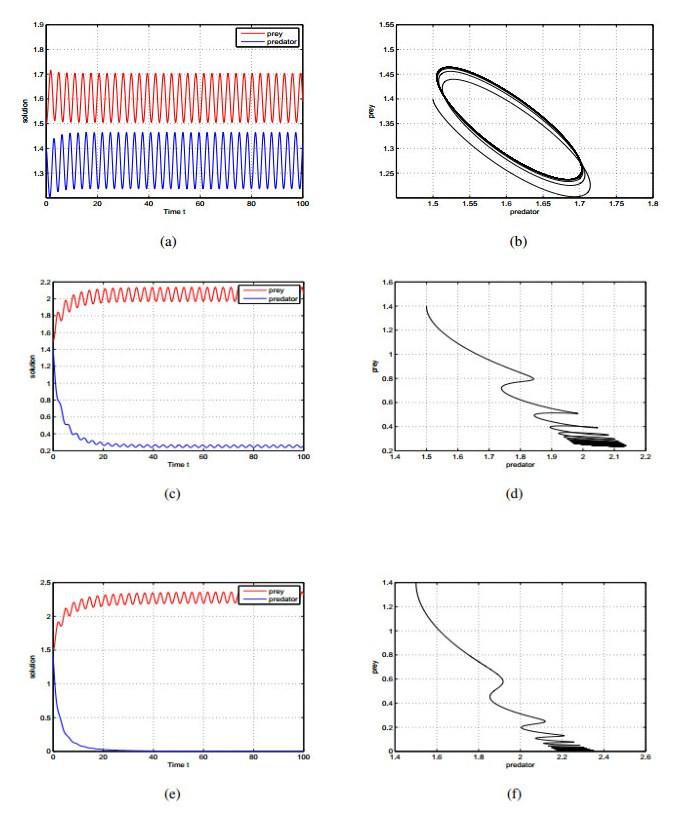
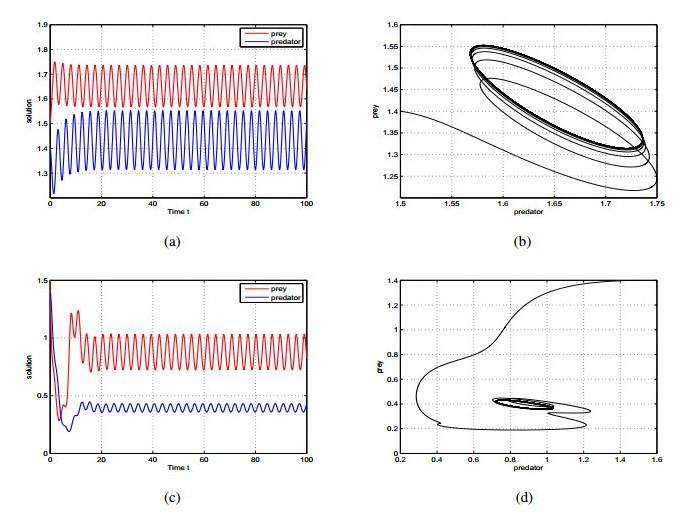
Figures(2)

Luoyi Wu, Hang Zheng, Songchuan Zhang. Dynamics of a non-autonomous predator-prey system with Hassell-Varley-Holling Ⅱ function response and mutual interference[J]. AIMS Mathematics, 2021, 6(6): 6033-6049. doi: 10.3934/math.2021355







 DownLoad:
DownLoad: