In this paper, the existence of positive periodic solution of stage-structured predator-prey model with Crowley-Martin type functional response is investigated. The prey population fall into two categories: mature and immature prey. The predator population is dependent only on mature prey and is influenced by Crowley-Martin type functional response. Based on the Mawhin's coincidence degree theory and nontrivial estimation techniques for a priori bounds of unknown solutions to the operator equation $ Fz = \mu Nz $, we prove the existence of positive periodic solution. Finally, the effectiveness of our result is verified by an example and numerical simulation.
Citation: Weijie Lu, Yonghui Xia. Periodic solution of a stage-structured predator-prey model with Crowley-Martin type functional response[J]. AIMS Mathematics, 2022, 7(5): 8162-8175. doi: 10.3934/math.2022454
In this paper, the existence of positive periodic solution of stage-structured predator-prey model with Crowley-Martin type functional response is investigated. The prey population fall into two categories: mature and immature prey. The predator population is dependent only on mature prey and is influenced by Crowley-Martin type functional response. Based on the Mawhin's coincidence degree theory and nontrivial estimation techniques for a priori bounds of unknown solutions to the operator equation $ Fz = \mu Nz $, we prove the existence of positive periodic solution. Finally, the effectiveness of our result is verified by an example and numerical simulation.

| [1] | A. Lotka, Elements of Physical Biology, USA: Williams Wilkins Co., Balitmore, 1925. |
| [2] | V. Volterra, Variazioni e fluttuazioni del numero dindividui in specie animali conviventi, Mem. Acad Lincei Roma., 2 (1926), 31–113. |
| [3] |
D. Wang, Positive periodic solutions for a nonautonomous neutral delay prey-predator model with impulse and Hassell-Varley type functional response, Proc. Amer. Math. Soc., 142 (2014), 623–638. https://doi.org/10.1090/S0002-9939-2013-11793-4 doi: 10.1090/S0002-9939-2013-11793-4

|
| [4] |
S. Yu, J. Liu, X. Lin, Multiple positive periodic solutions of a Gause-type predator-prey model with Allee effect and functional responses, AIMS Math., 5 (2020), 6135–6148. https://doi.org/10.3934/math.2020394 doi: 10.3934/math.2020394
|
| [5] | Y. Liu, X. Zhang, T. Zhou, Multiple periodic solutions of a delayed predator-prey model with non-monotonic functional response and stage structure, J. Biol. Dynam., 1 (2014), 145–160. |
| [6] |
S. Kant, V. Kumar, Stability analysis of predator-prey system with migrating prey and disease infection in both species, Appl. Math. Model., 42 (2017), 509–539. https://doi.org/10.1016/j.apm.2016.10.003 doi: 10.1016/j.apm.2016.10.003
|
| [7] |
S. Liu, L. Chen, Z. Liu, Extinction and permanence in nonautonomous competitive system with stage structure, J. Math. Anal. Appl., 274 (2002), 667–684. https://doi.org/10.1016/S0022-247X(02)00329-3 doi: 10.1016/S0022-247X(02)00329-3
|
| [8] | T. Zhang, H. Li, N. Xi, W. Fu, K. Wang, X. Ding, Mathematical analysis and simulation of a Hepatitis $B$ model with time delay: A case study for Xinjiang, China, Math. Biosci. Eng., 17 (2020), 1757–1775. |
| [9] |
Y. Song, X. Tang, Stability, steady-state bifurcations and turing patterns in a predator-prey model with herd behavior and prey-taxis, Stud. Appl. Math., 139 (2017), 371–404. https://doi.org/10.1111/sapm.12165 doi: 10.1111/sapm.12165
|
| [10] |
Y. Song, S. Wu, H. Wang, Spatiotemporal dynamics in the single population modelwith memory-based diffusion and nonlocal effect, J. Differential Equations, 267 (2019), 6316–6351. https://doi.org/10.1016/j.jde.2019.06.025 doi: 10.1016/j.jde.2019.06.025
|
| [11] |
X. Chen, Z. Du, Existence of positive periodic solutions for a neutral delay predator-prey model with Hassell-Varley type functional response and impulse, Qual. Theor. Dyn. Syst., 17 (2018), 67–80. https://doi.org/10.1007/s12346-017-0223-6 doi: 10.1007/s12346-017-0223-6
|
| [12] | Z. Wei, Y. Xia, T. Zhang, Stability and bifurcation analysis of a amensalism model with weak Allee effect, Qual. Theor. Dyn. Syst., 19 (2020), 23. |
| [13] | Z. Wei, Y. Xia, T. Zhang, Stability and bifurcation analysis of a commensal model with additive Allee effect and nonlinear growth rate, Int. J. Bifurcat. Chaos, 31 (2021), 2150204. |
| [14] |
R. Xu, Z. Ma, Stability and Hopf bifurcation in a ratio-dependent predator prey system with stage structure, Chaos Soliton. Fract., 38 (2008), 669–684. https://doi.org/10.1016/j.chaos.2007.01.019 doi: 10.1016/j.chaos.2007.01.019
|
| [15] |
J. Xu, T. Zhang, K. Song, A stochastic model of bacterial infection associated with neutrophils, Appl. Math. Comput., 373 (2020), 125025. https://doi.org/10.1016/j.amc.2019.125025 doi: 10.1016/j.amc.2019.125025
|
| [16] |
L. Chen, F. Chen, Qualitative analysis of a predator-prey model with Holling type Ⅱ functional response incorporating a constant prey refuge, Nonlinear Analysis, 11 (2010), 246–252. https://doi.org/10.1016/j.nonrwa.2008.10.056 doi: 10.1016/j.nonrwa.2008.10.056
|
| [17] |
F. Chen, Positive periodic solutions of neutral Lotka-Volterra system with feedback control, Appl. Math. Comput., 162 (2005), 1279–1302. https://doi.org/10.1016/j.amc.2004.03.009 doi: 10.1016/j.amc.2004.03.009
|
| [18] | F. Chen, On a nonlinear non-autonomous predator-prey model with diffusion and distributed delay, J. Comput. Appl. Math., 180 (2005), 33–49. |
| [19] | M. Agarwal, S. Devi, Persistence in a ratio-dependent predator-prey resource model with stage structure for prey, Int. J. Biomaths., 3 (2010), 313–336. |
| [20] |
S. Khajanchi, Modeling the dynamics of stage-structure predator-prey system with monod-haldane type functional response, Appl. Math. Comput., 302 (2017), 122–143. https://doi.org/10.1016/j.amc.2017.01.019 doi: 10.1016/j.amc.2017.01.019
|
| [21] |
M. Liu, K. Wang, Global stability of stage-structure predator-prey models with beddington de angelis functional response, Commun. Nonlinear Sci. Numer. Simul., 16 (2011), 3792–3797. https://doi.org/10.1016/j.cnsns.2010.12.026 doi: 10.1016/j.cnsns.2010.12.026
|
| [22] |
F. Chen, M. You, Permanence, extinction and periodic solution of the predator-prey system with Beddington-DeAngelis functional response and stage structure for prey, Nonlinear Anal., 9 (2008), 207–221. https://doi.org/10.1016/j.nonrwa.2006.09.009 doi: 10.1016/j.nonrwa.2006.09.009
|
| [23] |
C. Liu, Q. Zhang, L. Jinna, W. Yue, Stability analysis in a delayed prey-predator resource model with harvest effort and stage structure, Appl. Math. Comput., 238 (2014), 177–192. https://doi.org/10.1016/j.amc.2014.04.015 doi: 10.1016/j.amc.2014.04.015
|
| [24] | X. Ma, Y. Shao, Z. Wang, M. Luo, X. Fang, Z. Ju, An impulsive two-stage predator-prey model with stage structure and square root functional response, Math. Comput. Simulation, 119 (2016), 91–107. |
| [25] |
C. Holling, The functional response of predators to prey density and its role in mimicry and population regulation, Mem. Entomol. Soc. Canada, 45 (1965), 1–60. https://doi.org/10.4039/entm9745fv doi: 10.4039/entm9745fv
|
| [26] |
G. Skalski, J. Gilliam, Functional responses with predator interference: viable alternatives to the holling type-Ⅱ model, Ecological, 82 (2001), 3083–3092. https://doi.org/10.1890/0012-9658(2001)082[3083:FRWPIV]2.0.CO; 2 doi: 10.1890/0012-9658(2001)082[3083:FRWPIV]2.0.CO;2
|
| [27] |
J. Beddington, Mutual interference between parasites or predators and its effects on searching effiency, J. Anim. Ecol., 44 (1975), 331–340. https://doi.org/10.2307/43628162 doi: 10.2307/43628162
|
| [28] |
D. De-Angelis, R. Goldstein, R. O'Neill, A model for tropic interaction, Ecological, 56 (1975), 881–892. https://doi.org/10.2307/1936298 doi: 10.2307/1936298
|
| [29] |
X. Meng, H. Huo, H. Xiang, Q. Yin, Stability in a predator-prey model with Crowley-Martin function and stage structure for prey, Appl. Math. Comput., 232 (2014), 810–819. https://doi.org/10.1016/j.amc.2014.01.139 doi: 10.1016/j.amc.2014.01.139
|
| [30] | M. Cai, S. Yan, Z. Du, Positive periodic solutions of an Eco-Epidemic model with Crowley-Martin type functional response and disease in the Prey, Qual. Theor. Dyn. Syst., 19 (2020), 1–20. |
| [31] |
P. Crowley, E. Martin, Functional responses and interference within and between year classes of a dragonfly population, J. North. Am. Benth. Soc., 8 (1989), 211–221. https://doi.org/10.2307/1467324 doi: 10.2307/1467324
|
| [32] |
A. Maiti, B. Dubey, A. Chakraborty, Global analysis of a delayed stage structure prey-predator model with Crowley-Martin type functional response, Math. Comput. Simulation, 162 (2019), 58–84. https://doi.org/10.1016/j.matcom.2019.01.009 doi: 10.1016/j.matcom.2019.01.009
|
| [33] | A. Kosov, E. Semenov, On analytic periodic solutions to nonlinear differential equations with delay (Advance) Russ. Math., 62 (2018), 30–36. https://doi.org/10.3103/S1066369X18100043 |
| [34] |
A. Kosov, E. Semenov, First integrals and periodic solutions of a system with power nonlinearities, J. Appl. Ind. Math., 12 (2018), 70–83. https://doi.org/10.1134/S1990478918010076 doi: 10.1134/S1990478918010076
|
| [35] |
M. Fan, Q. Wang, X. Zou, Dynamics of a nonautonomous ratio-dependent predator-prey system, P. Roy. Soc. Lond. A. Math., 133 (2003), 97–118. https://doi.org/10.1023/A:1024156622489 doi: 10.1023/A:1024156622489
|
| [36] |
M. Fan, P. Wong, R. Agarwal, Periodicity and stability in periodic n-species Lotka-Volterra competition system with feedback controls and deviating arguments, Acta. Math. Sin., 19 (2003), 801–822. https://doi.org/10.1007/s10114-003-0311-1 doi: 10.1007/s10114-003-0311-1
|
| [37] | R. Gaines, J. Mawhin, Coincidence Degree and Nonlinear Differential Equations, NewYork: Springer-verlag, 1977. |
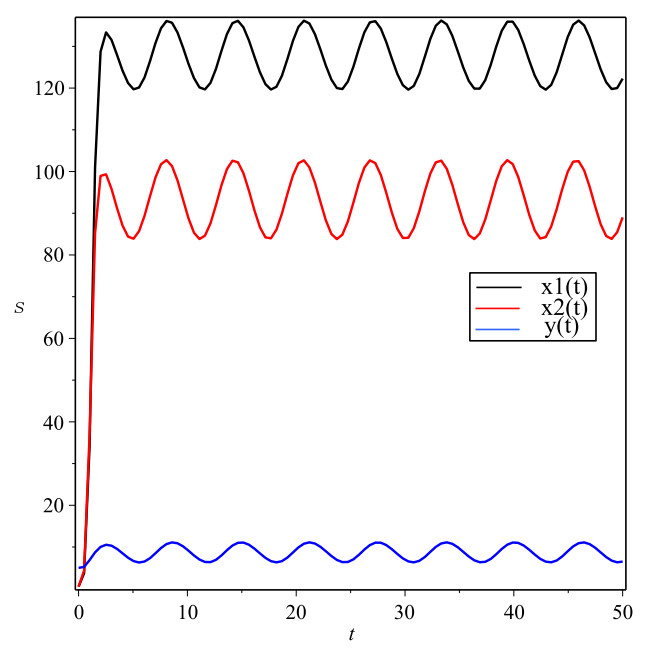
Figures(1)

Weijie Lu, Yonghui Xia. Periodic solution of a stage-structured predator-prey model with Crowley-Martin type functional response[J]. AIMS Mathematics, 2022, 7(5): 8162-8175. doi: 10.3934/math.2022454







 DownLoad:
DownLoad: