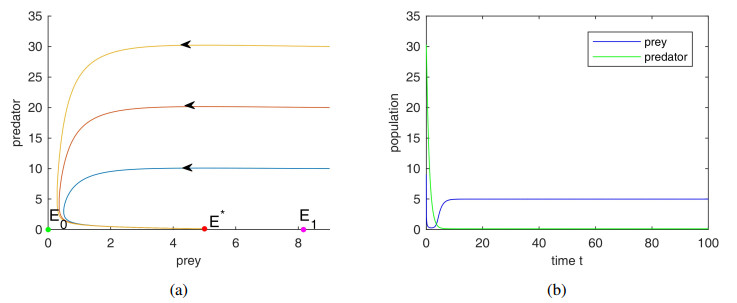
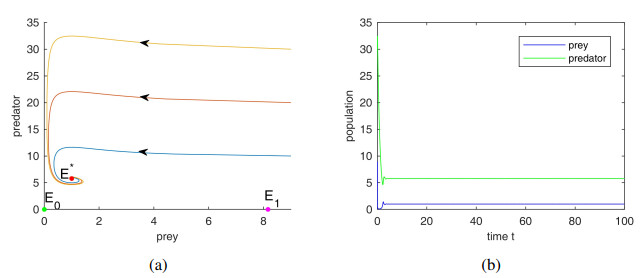
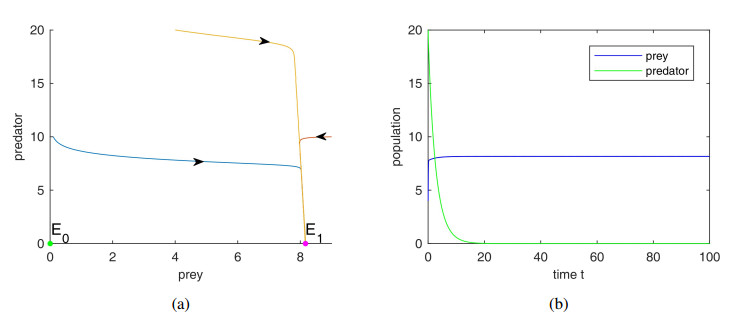
The aim of the paper is to study the impact of anti-predator behavior caused by dread of predator species in a prey predator system with Holling III type functional response and prey shelters. Firstly, we analyze the dynamic behavior of the system, including the stability of the system and demonstrating the occurrence of Hopf bifurcation around the positive equilibrium point and the existence of limit cycle emerging through Hopf bifurcation. Secondly, through the study of the effect of fear and refuge, we discover that the increase of fear level can improve the stability of the system by eliminating periodic solutions and decrease the populations of predator species at the coexist equilibrium, but not cause the extinction of the predators, and prey refuge also plays very vital role in the persistence of the predators. Finally, the rationality of the results is verified by numerical simulation.
Citation: Binfeng Xie, Na Zhang. Influence of fear effect on a Holling type III prey-predator system with the prey refuge[J]. AIMS Mathematics, 2022, 7(2): 1811-1830. doi: 10.3934/math.2022104
The aim of the paper is to study the impact of anti-predator behavior caused by dread of predator species in a prey predator system with Holling III type functional response and prey shelters. Firstly, we analyze the dynamic behavior of the system, including the stability of the system and demonstrating the occurrence of Hopf bifurcation around the positive equilibrium point and the existence of limit cycle emerging through Hopf bifurcation. Secondly, through the study of the effect of fear and refuge, we discover that the increase of fear level can improve the stability of the system by eliminating periodic solutions and decrease the populations of predator species at the coexist equilibrium, but not cause the extinction of the predators, and prey refuge also plays very vital role in the persistence of the predators. Finally, the rationality of the results is verified by numerical simulation.

| [1] |
K. Cheng, On the uniqueness of a limit cycle for a predator-prey system, SIAM J. Math. Anal., 12 (1988), 541–548. doi: 10.1137/0512047. doi: 10.1137/0512047

|
| [2] |
R. S. Cantrell, C. Cosner, On the dynamics of predator-prey models with the Beddington-DeAngelis functional response, J. Math. Anal. Appl., 257 (2001), 206–222. doi: 10.1006/jmaa.2000.7343. doi: 10.1006/jmaa.2000.7343
|
| [3] |
X. Liu, L. Chen, Complex dynamics of Holling II Lotka-Volterra predator-prey system with impulsive perturbations on the predator, Chaos Soliton. Fract., 16 (2003), 311–320. doi: 10.1016/S0960-0779(02)00408-3. doi: 10.1016/S0960-0779(02)00408-3
|
| [4] |
T. W. Hwang, Global analysis of the predator-prey system with Beddington-DeAngelis functional response, J. Math. Anal. Appl., 281 (2003), 395–401. doi: 10.1016/S0022-247X(02)00395-5. doi: 10.1016/S0022-247X(02)00395-5
|
| [5] |
X. Chen, F. Chen, Stable periodic solution of a discrete periodic Lotka-Volterra competition system with a feedback control, Appl. Math. Comput., 181 (2006), 1446–1454. doi: 10.1016/j.amc.2006.02.039. doi: 10.1016/j.amc.2006.02.039
|
| [6] |
F. Chen, Permanence and global attractivity of a discrete multispecies Lotka-Volterra competition predator-prey systems, Appl. Math. Comput., 182 (2006), 3–12. doi: 10.1016/j.amc.2006.03.026. doi: 10.1016/j.amc.2006.03.026
|
| [7] |
E. Gonzalez-Olivares, Multiple limit cycles in a gause type predator-prey model with Holling type III functional response and Allee effect on prey, Bull. Math. Biol., 73 (2011), 1378–1397. doi: 10.1007/s11538-010-9577-5. doi: 10.1007/s11538-010-9577-5
|
| [8] |
C. Shen, Permanence and global attractivity of the food-chain system with Holling IV type functional response, Appl. Math. Comput., 194 (2007), 179–185. doi: 10.1016/j.amc.2007.04.019. doi: 10.1016/j.amc.2007.04.019
|
| [9] |
N. Zhang, Y. Kao, F. Chen, B. Xie, S. Li, On a predator-prey system interaction under fluctuating water level with nonselective harvesting, Open Math., 18 (2020), 458–475. doi: 10.1515/math-2020-0145. doi: 10.1515/math-2020-0145
|
| [10] |
V. Volterra, Fluctuations in the abundance of a species considered mathematically, Nature, 118 (1926), 558–560. doi: 10.1038/118558a0. doi: 10.1038/118558a0
|
| [11] |
A. J. Lotka, Elements of physical biology, Am. J. Public Health, 21 (1926), 341–343. doi: 10.1038/116461b0. doi: 10.1038/116461b0
|
| [12] |
S. C. Holling, Some characteristics of simple types of predation and parasitism, Can. Entomol., 91 (1959), 385–398. doi: 10.4039/Ent91385-7. doi: 10.4039/Ent91385-7
|
| [13] |
T. Kar, H. Matsuda, Global dynamics and controllability of a harvested prey-predator system with Holling type III functional response, Nonlinear Anal. Hybrid Syst., 1 (2007), 59–67. doi: 10.1016/j.nahs.2006.03.002. doi: 10.1016/j.nahs.2006.03.002
|
| [14] |
W. Li, S. Wu, Traveling waves in a diffusive predator-prey model with Holling type-III functional response, Chaos Soliton. Fract., 37 (2008), 476–486. doi: 10.1016/j.chaos.2006.09.039. doi: 10.1016/j.chaos.2006.09.039
|
| [15] |
Y. Lamontagne, C. Coutu, C. Rousseau, Bifurcation analysis of a predator-prey system with generalised Holling type III functional response, J. Dyn. Differ. Equ., 20 (2008), 535–571. doi: 10.1007/s10884-008-9102-9. doi: 10.1007/s10884-008-9102-9
|
| [16] |
N. Apreutesei, G. Dimitriu, On a prey-predator reaction-diffusion system with Holling type III functional response, J. Comput. Appl. Math., 235 (2010), 366–379. doi: 10.1016/j.cam.2010.05.040. doi: 10.1016/j.cam.2010.05.040
|
| [17] |
J. Huang, S. Ruan, J. Song, Bifurcations in a predator-prey system of leslie type with generalized Holling type III functional response, J. Differ. Equations, 257 (2014), 1721–1752. doi: 10.1016/j.jde.2014.04.024. doi: 10.1016/j.jde.2014.04.024
|
| [18] |
R. Yang, J. Wei, Stability and bifurcation analysis of a diffusive prey-predator system in Holling type III with a prey refuge, Nonlinear Dynam., 79 (2015), 631–646. doi: 10.1007/s11071-014-1691-8. doi: 10.1007/s11071-014-1691-8
|
| [19] |
A. A. Shaikh, H. Das, N. Ali, Study of LG-Holling type III predator-prey model with disease in predator, J. Appl. Math. Comput., 58 (2018), 235–255. doi: 10.1007/s12190-017-1142-z. doi: 10.1007/s12190-017-1142-z
|
| [20] |
F. Rihan, C. Rajivganthi, Dynamics of fractional-order delay differential model of prey-predator system with Holling-type III and infection among predators, Chaos Soliton. Fract., 141 (2020), 110365. doi: 10.1016/j.chaos.2020.110365. doi: 10.1016/j.chaos.2020.110365
|
| [21] |
Y. Xie, Z. Wang, B. Meng, X. Huang, Dynamical analysis for a fractional-order prey-predator model with Holling III type functional response and discontinuous harvest, Appl. Math. Lett., 106 (2020), 106342. doi: 10.1016/j.aml.2020.106342. doi: 10.1016/j.aml.2020.106342
|
| [22] |
J. B. Collings, Bifurcation and stability analysis of a temperature-dependent mite predator-prey interaction model incorporating a prey refuge, Bull. Math. Biol., 57 (1995), 63–76. doi: 10.1007/BF02458316. doi: 10.1007/BF02458316
|
| [23] |
T. K. Kar, Stability analysis of a prey-predator model incorporating a prey refuge, Commun. Nonlinear Sci., 10 (2005), 681–691. doi: 10.1016/j.cnsns.2003.08.006. doi: 10.1016/j.cnsns.2003.08.006
|
| [24] |
T. K. Kar, Modelling and analysis of a harvested prey-predator system incorporating a prey refuge, J. Comput. Appl. Math., 185 (2006), 19–33. doi: 10.1016/j.cam.20050.01.035. doi: 10.1016/j.cam.20050.01.035
|
| [25] |
Y. Huang, F. Chen, L. Zhong, Stability analysis of a prey-predator model with Holling type III response function incorporating a prey refuge, Appl. Math. Comput., 182 (2006), 672–683. doi: 10.1016/j.amc.2006.04.030. doi: 10.1016/j.amc.2006.04.030
|
| [26] |
W. Ko, K. Ryu, Qualitative analysis of a predator-prey model with Holling type II functional response incorporating a prey refuge, J. Differ. Equations, 231 (2006), 534–550. doi: 10.1016/j.jde.2006.08.001. doi: 10.1016/j.jde.2006.08.001
|
| [27] |
F. Chen, Z. Ma, H. Zhang, Global asymptotical stability of the positive equilibrium of the Lotka-Volterra preypredator model incorporating a constant number of prey refuges, Nonlinear Anal-Real., 13 (2012), 2790–2793. doi: 10.1016/j.nonrwa.2012.04.006. doi: 10.1016/j.nonrwa.2012.04.006
|
| [28] |
Y. Wang, J. Wang, Influence of prey refuge on predator-prey dynamics, Nonlinear Dynam., 67 (2012), 191–201. doi: 10.1007/s11071-011-9971-z. doi: 10.1007/s11071-011-9971-z
|
| [29] |
J. P. Tripathi, S. Abbas, M. Thakur, Dynamical analysis of a prey-predator model with Beddington-DeAngelis type function response incorporating a prey refuge, Nonlinear Dynam., 80 (2015), 177–196. doi: 10.1007/s11071-014-1859-2. doi: 10.1007/s11071-014-1859-2
|
| [30] |
R. Yang, J. Wei, Stability and bifurcation analysis of a diffusive prey-predator system in Holling type III with a prey refuge, Nonlinear Dynam., 79 (2015), 631–646. doi: 10.1007/s11071-014-1691-8. doi: 10.1007/s11071-014-1691-8
|
| [31] |
S. Banerjee, S. Khajanchi, Role of constant prey refuge on stage structure predator-prey model with ratio dependent functional response, Appl. Math. Comput., 314 (2017), 193–198. doi: 10.1016/j.amc.2017.07.017. doi: 10.1016/j.amc.2017.07.017
|
| [32] |
Y. Wu, F. Chen, C. Du, Dynamic behaviors of a nonautonomous predator-prey system with Holling type II schemes and a prey refuge, Adv. Differ. Equ., 2021 (2021), 62. doi: 10.1186/s13662-021-03222-1. doi: 10.1186/s13662-021-03222-1
|
| [33] |
X. Wang, L. Zanette, X. Zou, Modelling the fear effect in predator-prey interactions, J. Math. Biol., 73 (2016), 1179–1204. doi: 10.1007/s00285-016-0989-1. doi: 10.1007/s00285-016-0989-1
|
| [34] |
H. Zhang, Y. Cai, S. Fu, W. Wang, Impact of the fear effect in a prey-predator model incorporating a prey refuge, Appl. Math. Comput., 356 (2019), 328–337. doi: 10.1016/j.amc.2019.03.034. doi: 10.1016/j.amc.2019.03.034
|
| [35] |
K. Sarkar, S. Khajanchi, Impact of fear effect on the growth of prey in a predator-prey interaction model, Ecol. Complex., 42 (2020), 100826. doi: 10.1016/j.ecocom.2020.100826. doi: 10.1016/j.ecocom.2020.100826
|
| [36] |
L. Lai, Z. Zhu, F. Chen, Stability and bifurcation in a predator-prey model with the additive Allee effect and the fear effect, Mathematics, 8 (2020), 1280. doi: 10.3390/math8081280. doi: 10.3390/math8081280
|
| [37] |
R. P. Kaur, A. Sharma, A. K. Sharma, Impact of fear effect on plankton-fish system dynamics incorporating zooplankton refuge, Chaos Soliton. Fract., 143 (2021), 110563. doi: 10.1016/j.chaos.2020.110563. doi: 10.1016/j.chaos.2020.110563
|
| [38] |
B. Xie, Z. Zhang, N. Zhang, Influence of the fear effect on a Holling Type II prey-predator system with a Michaelis-Menten type harvesting, Int. J. Bifurcat. Chaos, 31 (2021), 2150216. doi: 10.1142/S0218127421502163. doi: 10.1142/S0218127421502163
|
| [39] |
B. Xie, Impact of the fear and Allee effect on a Holling type II prey-predator model, Adv. Differ. Equ., 2021 (2021), 464. doi: 10.1186/s13662-021-03592-6. doi: 10.1186/s13662-021-03592-6
|
| [40] |
L. Y. Zanette, A. F. White, M. C. Allen, M. Clinchy, Perceived predation risk reduces the number of offspring songbirds produce per year, Science, 334 (2011), 1398–1401. doi: 10.1126/science.1210908. doi: 10.1126/science.1210908
|
| [41] |
S. K. Sasmal, Population dynamics with multiple allee effects induced by fear factors–A mathematical study on prey-predator interactions, Appl. Math. Model., 64 (2018), 1–14. doi: 10.1016/j.apm.2018.07.021. doi: 10.1016/j.apm.2018.07.021
|
| [42] |
X. Wang, X. Zou, Modeling the fear effect in predator-prey interactions with adaptive avoidance of predators, Bull. Math. Biol., 79 (2017), 1–35. doi: 10.1007/s11538-017-0287-0. doi: 10.1007/s11538-017-0287-0
|
| [43] | T. Burton, Volterra Integral and Differential Equations, Orlando: Academic Press, 1983. doi: 10.1137/1027032. |
| [44] | J. D. Meiss, Differential Dynamical Systems, Philadelphia: Society for Industrial and Applied Mathematics, 2007. |
| [45] |
J. Chen, H. Zhang, The qualitative analysis of two species predator-prey model with Holling's type III functional response, Appl. Math. Mech., 71 (1986), 73–80. doi: 10.1007/BF01896254. doi: 10.1007/BF01896254
|






Figures(13) / Tables(2)

Binfeng Xie, Na Zhang. Influence of fear effect on a Holling type III prey-predator system with the prey refuge[J]. AIMS Mathematics, 2022, 7(2): 1811-1830. doi: 10.3934/math.2022104







 DownLoad:
DownLoad: