Citation: Foday Conteh, Shota Tobaru, Mohamed E. Lotfy, Atsushi Yona, Tomonobu Senjyu. An effective Load shedding technique for micro-grids using artificial neural network and adaptive neuro-fuzzy inference system[J]. AIMS Energy, 2017, 5(5): 814-837. doi: 10.3934/energy.2017.5.814

| [1] |
Bakar NNA, Hassan MY, Sulaima MF, et al. (2017) Microgrid and load shedding scheme during islanded mode: A review. Renew S ust Energ Rev 71: 161-169. doi: 10.1016/j.rser.2016.12.049

|
| [2] |
Laghari JA, Mokhlis H, Bakar AHA, et al. (2013) Application of computational intelligence techniques for load shedding in power systems. Energy Convers Manage 75: 130-140. doi: 10.1016/j.enconman.2013.06.010
|
| [3] | Pushpanjali M, Sujatha MS (2015) A novel multi objective under frequency load shedding in a micro-grid using genetic algorithm. Int J Advan Res Electri Electro Instru Eng 4: 5037-5038. |
| [4] | Kaffashan I, Mortezaee SMTM, Amraee T (2016) A robust under voltage load shedding scheme against voltage. Turk J Electr Eng Comp S ci 24: 3310-3311. |
| [5] |
Hooshmand R, Moazzami M (2012) Optimal design of adaptive under frequency load shedding using artificial neural networks in isolated power system. Int J Elec Power 42: 220-228. doi: 10.1016/j.ijepes.2012.04.021
|
| [6] | Boland J, Burdett R, Edwards C (2015) Optimization techniques for planning automatic under frequency load shedding in New Zealand's power system. Anziam J 57: M3-M4. |
| [7] | Suwanasri C, Suwanasri T, Prachuab N (2013) Load shedding control strategy in an electric distribution system. GMS RAN Int J 7: 47-52. |
| [8] | Moazzami M, Morshed MJ, Fekih A, et al. (2016) A new optimal unified power flow controller placement and load shedding coordination approach using the hybrid imperialist competitive algorithm-pattern search method for voltage collapse prevention in power system Int J Elec Power 79: 263-274. |
| [9] |
Aponte EE, Nelson JK (2006) Time optimal shedding for distributed power systems. IEEE T Power S yst 21: 269-277. doi: 10.1109/TPWRS.2005.857826
|
| [10] | Das K, Nitsas A, Altin M, et al. (2016) Improved load shedding scheme considering distributed generation. IEEE T Power Delivery, 1-4. |
| [11] | Malekpour AR, Seifi AR, Hesamzadeh MR, et al. (2008) An optimal load shedding approach for distribution networks with DGs considering capacity deficiency modelling of bulked power supply. World Academic S ci Eng Tech 22: 825-826. |
| [12] |
Marzband M, Moghaddm MM, Akorede MF, et al. (2016) Adaptive load shedding scheme for frequency stability enhancement in micro-grids. Electr Pow S yst Res 140: 78-86. doi: 10.1016/j.epsr.2016.06.037
|
| [13] |
Tahir MF, Tehzeeb-ul-Hassan, Saqib MA (2016) Optimal scheduling of electrical power in energy-deficiency scenarios using artificial neural network and bootstrap aggregation. Int J Elec Power 83: 49-53. doi: 10.1016/j.ijepes.2016.03.046
|
| [14] | Kuriakose E, Francis F (2013) Enhancement of power system stability by optimal adaptive under frequency load shedding using artificial neural networks. Int J Advan Res Electri Electro Instru Eng 2: 12-20. |
| [15] |
Amooshahi H, Rahmat-Allah H, Khodabakhshian A, et al. (2016) A new load-shedding approach for micro-grids in the presence of wind turbines. Electr Pow Compo S ys 44: 1-11. doi: 10.1080/15325008.2015.1091861
|
| [16] |
Koohi-Kamali S, Rahim NA (2016) Coordinated control of smart micro-grid during and after is landing operation to prevent under frequency load shedding using energy storage system. Energ Convers Manage 127: 623-646. doi: 10.1016/j.enconman.2016.09.052
|
| [17] |
Laghari JA, Mokhlis H, Karimi M, et al. (2015) A new under-frequency load shedding technique based on combination of fixed and random priority of loads for smart grid applications. IEEE T Power S yst 30: 2507-2515. doi: 10.1109/TPWRS.2014.2360520
|
| [18] |
Sapari NM, Mokhlis H, Laghari JA, et al. (2017) Load shedding scheme based on frequency and voltage stability for islanding operation of distribution network connected to mini-hydro generation. Turk J Electr Eng Co 25: 1852-1863. doi: 10.3906/elk-1601-111
|
| [19] |
Karimi M, Mohamad H, Mokhlis H, et al. (2012) Under-Frequency load Shedding scheme for islanded distribution network connected to mini-hydro generation. Electrical Power Energy S ystems 42: 127-138. doi: 10.1016/j.ijepes.2012.03.017
|
| [20] |
Khezri R, Golshannavaz S, Vakili R, et al. (2017) Multi-layer fuzzy-based under-frequency load shedding in back-pressure smart industrial microgrids. Energy 132: 96-105. doi: 10.1016/j.energy.2017.05.059
|
| [21] | Dragomir OE, Dragomir F, Stefan V, et al. (2014) Adaptive neuro-fuzzy inference systems as a strategy for predicting and controlling the energy produced from renewable sources. Energies 8: 13047-13061. |
| [22] | Bhatnagar S, Chopra S, Bhati S, et al. (2015) Under frequency load shedding using adaptive neuro-fuzzy inference system. IJEEE 7: 99. |
| [23] |
Lakra P, Kirar M (2015) Load shedding techniques for system with cogeneration: A review. ELELIJ 4: 83-96. doi: 10.14810/elelij.2015.4307
|
| [24] | Khamis A, Shareef H, Mohamed A, et al. (2015) A load shedding scheme for DG integrated islanded power system utilizing backtracking search algorithm. Ain S hams Eng J, 2-3. |
| [25] | Swetha GC, Sudarshana RHR (2017) Voltage stability assessment in power network using artificial neural network. Int J Advan Res Electri Electro and Instru Eng, 7993-8002. |
| [26] | Kuriakose E, Francis F (2017) Enhancement of power system stability by optimal adaptive under frequency load shedding using artificial neural networks. Int J Advan Res Electri Electro Instru Eng, 12-20. |
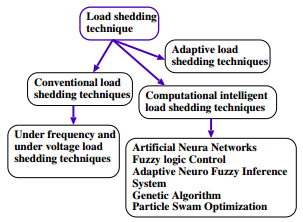
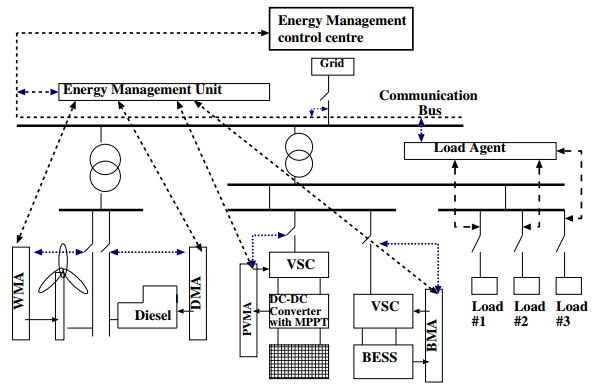
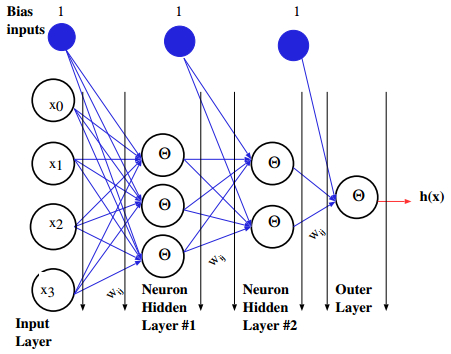
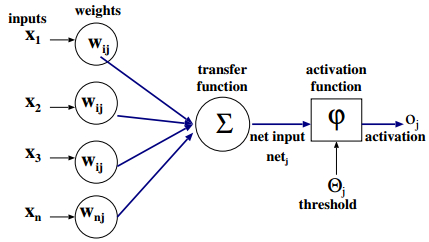
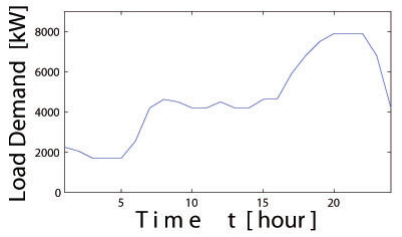

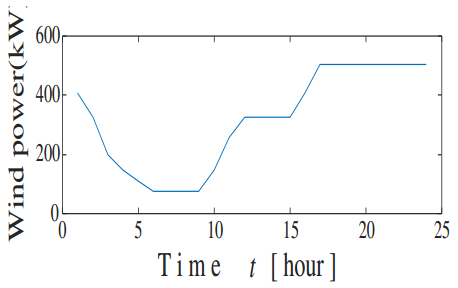
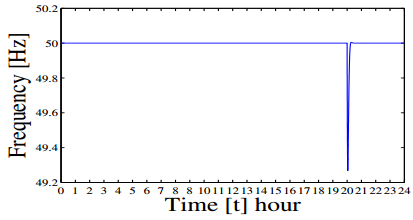
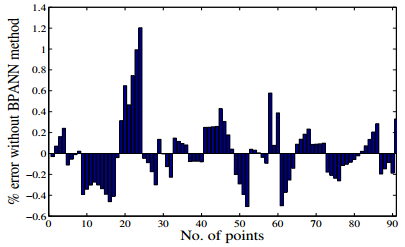


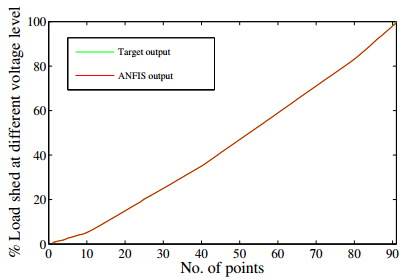
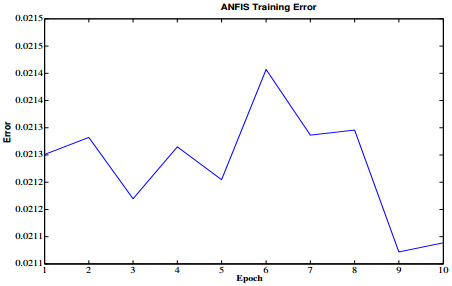
Figures(21) / Tables(3)

Foday Conteh, Shota Tobaru, Mohamed E. Lotfy, Atsushi Yona, Tomonobu Senjyu. An effective Load shedding technique for micro-grids using artificial neural network and adaptive neuro-fuzzy inference system[J]. AIMS Energy, 2017, 5(5): 814-837. doi: 10.3934/energy.2017.5.814







 DownLoad:
DownLoad: