Citation: Natalia Fullana, Victoria Braña, Juan José Marizcurrena, Danilo Morales, Jean-Michel Betton, Mónica Marín, Susana Castro-Sowinski. Identification, recombinant production and partial biochemical characterization of an extracellular cold-active serine-metalloprotease from an Antarctic Pseudomonas isolate[J]. AIMS Bioengineering, 2017, 4(3): 386-401. doi: 10.3934/bioeng.2017.3.386

| [1] |
Wu JW, Chen XL (2011) Extracellular metalloproteases from bacteria. Appl Microbiol Biot 92: 253–262. doi: 10.1007/s00253-011-3532-8

|
| [2] | Rao MB, Tanksale AM, Ghatge MS, et al. (1998) Molecular and biotechnological aspects of microbial proteases. Microbiol Mol Biol Rev 62: 597–635. |
| [3] |
Kasana RC (2010) Proteases from psychrotrophs: an overview. Crit Rev Microbiol 36: 134–145. doi: 10.3109/10408410903485525
|
| [4] | Roulling F, Piette F, Cipolla A, et al. (2011) Psychrophilic enzymes: cool responses to chilly problems. In: Extremophiles Handbook. Koki Horikoshi (Ed.) Springer, 891–913. |
| [5] |
Joshi S, Satyanarayana T (2013) Biotechnology of cold-active proteases. Biology 2: 755–783. doi: 10.3390/biology2020755
|
| [6] | Martínezrosales C, Castrosowinski S (2011) Antarctic bacterial isolates that produce cold-active extracellular proteases at low temperature but are active and stable at high temperature. Polar Res 30: 157–171. |
| [7] |
Studier FW (2005) Protein production by auto-induction in high-density shaking cultures. Protein Expres Pur 41: 207–234. doi: 10.1016/j.pep.2005.01.016
|
| [8] |
Piñeyro MD, Parodi-Talice A, Portela M, et al. (2011) Molecular characterization and interactome analysis of Trypanosoma cruzi Tryparedoxin 1. J Proteomics 74: 1683–1692. doi: 10.1016/j.jprot.2011.04.006
|
| [9] | Morel MA, Iriarte A, Jara E, et al. (2016) Revealing the biotechnological potential of Delftia sp. JD2 by a genomic approach. AIMS Bioeng 3: 156–175. |
| [10] |
Aziz RK, Bartels D, Best AA, et al. (2008) The RAST server: rapid annotations using subsystems technology. BMC Genomics 9: 75. doi: 10.1186/1471-2164-9-75
|
| [11] |
Andrews BA, Asenjo JA (1986) Synthesis and regulation of extracellular β(1-3) glucanase and protease by Cytophaga sp. in batch and continuous culture. Biotechnol Bioeng 28: 1366–1375. doi: 10.1002/bit.260280911
|
| [12] |
D´Amico S, Claverie P, Collins T, et al. (2002) Molecular basis of cold adaptation. Philos T R Soc B 357: 917–925. doi: 10.1098/rstb.2002.1105
|
| [13] |
Martinez-Rosales C, Fullana N, Musto H, et al. (2012) Antarctic DNA moving forward: genomic plasticity and biotechnological potential. FEMS Microbiol Lett 331: 1–9. doi: 10.1111/j.1574-6968.2012.02531.x
|
| [14] |
De Vendittis E, Castellano I, Cotugno R, et al. (2008) Adaptation of model proteins from cold to hot environments involves continuous and small adjustments of average parameters related to amino acid composition. J Theor Biol 250: 156–171. doi: 10.1016/j.jtbi.2007.09.006
|
| [15] | Herrera LM, García-Laviña CX, Marizcurrena JJ, et al. (2016) Hydrolytic enzyme-producing microbes in the Antarctic oligochaete Grania sp. (Annelida). Polar Biol 40: 947–953. |
| [16] |
Zhou MY, Wang GL, Li D, et al. (2013) Diversity of both the cultivable protease-producing bacteria and bacterial extracellular proteases in the coastal sediments of King George Island, Antarctica. PLoS One 8: e79668. doi: 10.1371/journal.pone.0079668
|
| [17] | Martínez-Rosales C, Marizcurrena JJ, Iriarte A, et al. (2015) Characterizing proteases in an Antarctic Janthinobacterium sp. isolate: evidence of a protease horizontal gene transfer event. Adv Polar Sci 26: 88–95. |
| [18] | Lenders MH, Reimann S, Smits SH, et al. (2013) Molecular insights into type I secretion systems. Biol Chem 394: 1371–1384. |
| [19] |
De Maayer P, Anderson D, Cary C, et al. (2014) Some like it cold: understanding the survival strategies of psychrophiles. EMBO Rep 15: 508. doi: 10.1002/embr.201338170
|
| [20] |
Chessa JP, Petrescu I, Bentahir M, et al. (2000) Purification, physic-chemical characterization and sequence of a heat labile alkaline metalloprotease isolated from a psychrophilic Pseudomonas species. Biochim Biophys Acta 1479: 265–274. doi: 10.1016/S0167-4838(00)00018-2
|
| [21] |
Nicodeme M, Grill JP, Humbert G, et al. (2005) Extracellular protease activity of different Pseudomonas strains: dependence of proteolytic activity on culture conditions. J Appl Microbiol 99: 641–648. doi: 10.1111/j.1365-2672.2005.02634.x
|
| [22] | Dufour D, Nicodeme M, Perrin C, et al. (2008) Molecular typing of industrial strains of Pseudomonas spp. isolated from milk and genetical and biochemical characterization of an extracellular protease produced by one of them. Int J Food Microbiol 125: 188–196. |
| [23] | Liao CH, MaCcallus DE (1998) Biochemical and genetic characterization of an extracellular protease from Pseudomonas fluorescens CY 091. Appl Environ Microb 64: 914–921. |
| [24] | Ahn JH, Pan JG, Rhee JS (1999) Identification of the tliDEFABC transporter specific for lipase in Pseudomonas fluorescens SIK W1. J Bacteriol 181: 1847–1852. |
| [25] |
Kawai E, Idei A, Kumura H, et al. (1999) The ABC exporter genes involved in the lipase secretion are clustered with the genes for lipase, alkaline protease, and serine protease homologues in Pseudomonas fluorescens. Biochim Biophys Acta 1446: 377–382. doi: 10.1016/S0167-4781(99)00094-9
|
| [26] |
Hege T, Feltzer RE, Gray RD, et al. (2001) Crystal structure of a complex between Pseudomonas aeruginosa alkaline protease and its cognate inhibitor: Inhibition by a zinc-NH2 coordinative bond. J Biol Chem 276: 35087–35092. doi: 10.1074/jbc.M104020200
|
| [27] |
Speed MA, Wang DIC, King J (1996) Specific aggregation of partially folded polypeptide chains: the molecular basis of inclusion body composition. Nat Biotechnol 14: 1283–1287. doi: 10.1038/nbt1096-1283
|
| [28] | Ertan H, Cassel C, Verma A, et al. (2015) A new broad specificity alkaline metalloprotease from a Pseudomonas sp. isolated from refrigerated milk: Role of calcium in improving enzyme productivity. J Mol Catal B Enzym 113: 1–8. |
| [29] |
Wu S, Liu G, Zhang D, et al. (2015) Purification and biochemical characterization of an alkaline protease from marine bacteria Pseudoalteromonas sp. 129-1. J Basic Microb 55: 1427–1434. doi: 10.1002/jobm.201500327
|
| [30] | Niyonzima FN, More SS (2015) Purification and characterization of detergent-compatible protease from Aspergillus terreus gr. Biotech 5: 61–70. |
| [31] |
Raval VH, Pillai S, Rawal CM, et al. (2014) Biochemical and structural characterization of a detergent-stable serine alkaline protease from seawater haloalkaliphilic bacteria. Process Biochem 49: 955–962. doi: 10.1016/j.procbio.2014.03.014
|
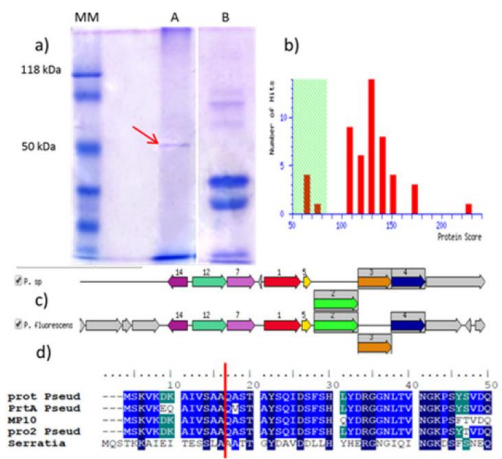

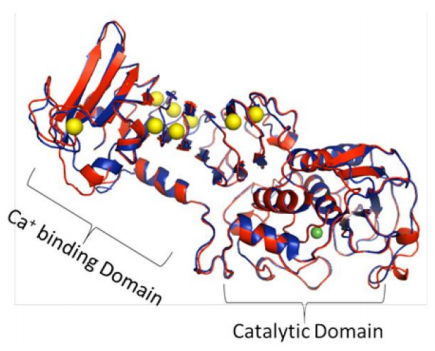
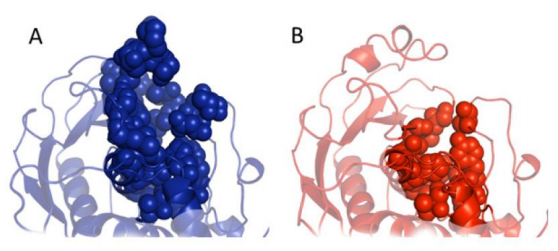
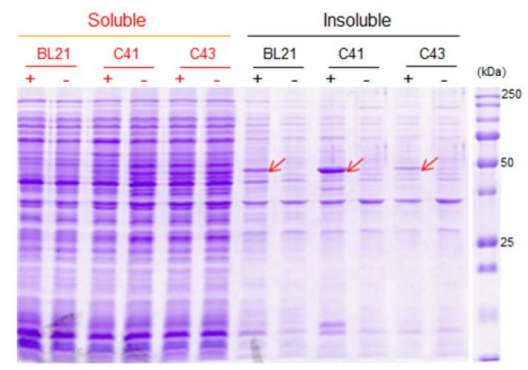
Figures(6) / Tables(2)

Natalia Fullana, Victoria Braña, Juan José Marizcurrena, Danilo Morales, Jean-Michel Betton, Mónica Marín, Susana Castro-Sowinski. Identification, recombinant production and partial biochemical characterization of an extracellular cold-active serine-metalloprotease from an Antarctic Pseudomonas isolate[J]. AIMS Bioengineering, 2017, 4(3): 386-401. doi: 10.3934/bioeng.2017.3.386







 DownLoad:
DownLoad: