Citation: Yang Li, Jia Li. Stage-structured discrete-time models for interacting wild and sterile mosquitoes with beverton-holt survivability[J]. Mathematical Biosciences and Engineering, 2019, 16(2): 572-602. doi: 10.3934/mbe.2019028

| [1] | L. Alphey, M. Benedict, R. Bellini, G. G. Clark, D. A. Dame, M. W. Service and S. L. Dobson, Sterile-insect methods for control of mosquito-borne diseases: An analysis, Vector-Borne Zoonot., 10 (2010), 295–311. |
| [2] | AMCA, Life Cycle. Available from: http://www.mosquito.org/page/lifecycle. |
| [3] | B. Anderson, J. Jackson and M. Sitharam, Descartes's rule of signs revisited, Amer. Math. Monthly 105 (1998), 447–451. |
| [4] | H. J. Barclay, The sterile insect release method for species with two-stage life cycles, Res. Popul. Ecol., 21 (1980), 165–180. |
| [5] | H. J. Barclay, Pest population stability under sterile releases, Res. Popul. Ecol., 24 (1982), 405– 416. |
| [6] | H. J. Barclay, Mathematical models for the use of sterile insects, in Sterile Insect Technique, Principles and Practice in Area-Wide Integrated Pest Management, V. A. Dyck, J. Hendrichs, and A. S. Robinson , (Eds. Springer, Heidelberg), (2005), 147–174. |
| [7] | H. J. Barclay and M. Mackuer, The sterile insect release method for pest control: a density dependent model, Environ. Entomol., 9 (1980), 810–817. |
| [8] | A. C. Bartlett and R. T. Staten, Sterile Insect Release Method and other Genetic Control Strategies, Radcliffe's IPM World Textbook, 1996, available from: http://ipmworld.umn.edu/chapters/bartlett.htm. |
| [9] | N. Becker, Mosquitoes and Their Control, Kluwer Academic/Plenum, New York, 2003. |
| [10] | M. Q. Benedict and A. S. Robinson, The first releases of transgenic mosquitoes: an argument for the sterile insect technique, Trends Parasitol., 19 (2003), 349–55. |
| [11] | R. J. H. Beverton and S. J. Holt, On the Dynamics of Exploited Fish Populations, Springer Science and Business Media, Jun 30, 1993. |
| [12] | M. Bohner and H. Warth, The Beverton-Holt dynamic equation, Appl. Anal., 86 (2007), 1007– 1015. |
| [13] | L. M. Cai, S. B. Ai and J. Li, Dynamics of mosquitoes populations with different strategies for releasing sterile mosquitoes, SIAM, J. Appl. Math., 74 (2014), 1786–1809. |
| [14] | J. M. Cushing and J. Li, On Ebenman's model for the dynamics of a population with competing juveniles and adults, Bull. Math. Biol., 51 (1989), 687–713. |
| [15] | C. Dye, Intraspecific competition amongst larval aedes aegypti: Food exploitation or chemical interference, Ecol. Entomol., 7 (1982), 39–46. |
| [16] | S. Elaydi, Discrete Chaos: With Applications in Science and Engineering, 2nd edition, Boca Raton, CRC Press, 2007. |
| [17] | K. R. Fister, M. L. McCarthy, S. F. Oppenheimer and Craig Collins, Optimal control of insects through sterile insect release and habitat modification, Math. Biosci., 244 (2013), 201–212. |
| [18] | J. C. Floresa, A mathematical model for wild and sterile species in competition: immigration, Physica A, 328 (2003), 214–224. |
| [19] | R. M. Gleiser, J. Urrutia and D. E. Gorla, Effects of crowding on populations of aedes albifasciatus larvae under laboratory conditions, Entomol. Exp. Appl., 95 (2000), 135–140. |
| [20] | D. J. Grabiner, Descartes's rule of signs: Another construction, Amer. Math. Monthly, 106 (1999), 854–855. |
| [21] | C. S. Holling, The functional response of predator to prey density and its role in mimicry and population regulation, Mem. Entomol. Soc. Can., 45 (1965), 5–60. |
| [22] | E. I. Jury, The inners approach to some problems of system theory, IEEE, Trans. Automatic Contr., AC-16 (1971), 233–240. |
| [23] | E. S. Krafsur, Sterile insect technique for suppressing and eradicating insect populations: 55 years and counting, J. Agr. Entomol., 15 (1998), 303–317. |
| [24] | J. Li, Simple stage-structured models for wild and transgenic mosquito populations, J. Diff. Eqns. Appl., 17 (2009), 327–347. |
| [25] | J. Li, Modeling of mosquitoes with dominant or recessive transgenes and Allee effects, Math. Biosci. Eng., 7 (2010), 101–123. |
| [26] | J. Li, Malaria model with stage-structured mosquitoes, Math. Biol. Eng., 8 (2011), 753–768. |
| [27] | J. Li, Stage-structured models for interacting wild and sterile mosquitoes, Journal of Shanghai Normal University, 43 (2014), 511–522. |
| [28] | J. Li, New revised simple models for interactive wild and sterile mosquito populations and their dynamics, J. Biol. Dynam., 11(S2) (2017). 316–333. |
| [29] | J. Li and Z. L. Yuan, Modeling releases of sterile mosquitoes with different strategies, J. Biol. Dynam., 9 (2015), 1–14. |
| [30] | Y. Li, Discrete-time Structured Models and their Dynamics for Interactive Wild and Sterile Mosquitoes and Malaria Transmissions, Ph.D thesis, The University of Alabama in Huntsville, 2017. |
| [31] | Y. Li and J. Li, Discrete-time models for releases of sterile mosquitoes with Beverton-Holt-type of survivability, Ricerche di Matematica, 67 (2018), 141–162. |
| [32] | Y. Li and J. Li, Discrete-time model for malaria transmission with constant releases of sterile mosquitoes, preprint, 2018. |
| [33] | R. M. May, G. R. Conway, M. P. Hassell and T. R. E. Southwood, Time delays, density-dependence and single-species oscillations, J. Anim. Ecol., 43 (1974), 747–770. |
| [34] | Mosquito Facts, 33 Things You Didn't Know About Mosquitoes, https://www.megacatch. com/mosquito-faqs/mosquito-facts/. |
| [35] | M. Otero, H. G. Solari and N. Schweigmann, A stochastic population dynamics model for Aedes aegypti: formulation and application to a city with temperate climate, Bull. Math. Biol., 68 (2006), 1945–1974. |
| [36] | D. Ruiz, G. Poveda, I. D. Velez, M. L. Quinones, G. L. Rua, L. E.Velasquesz and J. S. Zuluage, Modeling entomological-climatic interactions of Plasmodium falciparum malaria transmission in two Colombian endemic-regions: contributions to a National Malaria Early Warning System, Malaria J., 5 (2006), 66. |
| [37] | Wikipedia, Mosquito-borne disease. Available from: https://en.wikipedia.org/wiki/ Mosquito-borne_disease. |
| [38] | Wikipedia, Sterile insect technique. Available from: https://en.wikipedia.org/wiki/ Sterile_insect_technique. |
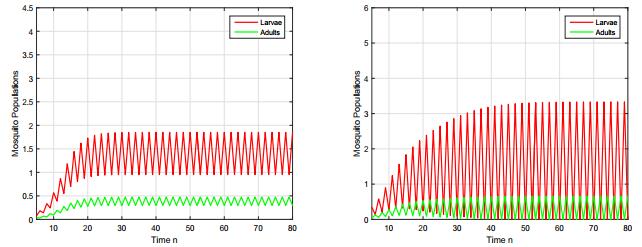
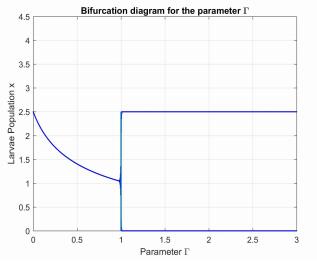
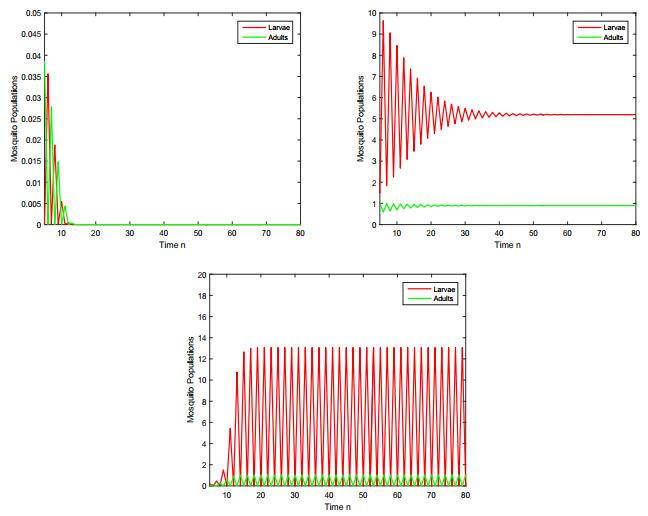
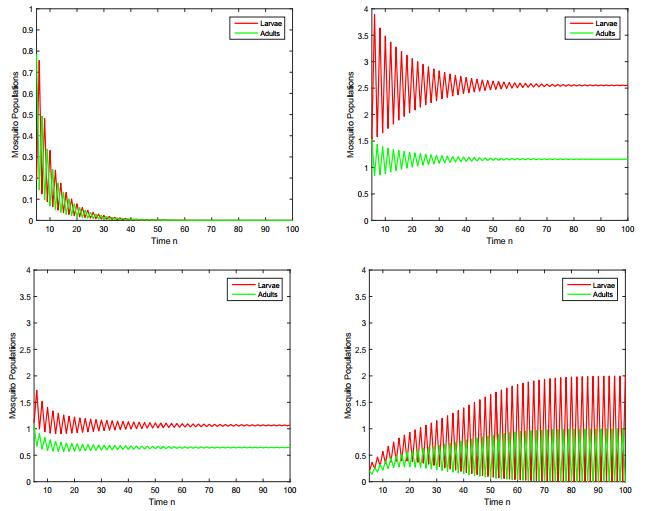
Figures(8) / Tables(3)

Yang Li, Jia Li. Stage-structured discrete-time models for interacting wild and sterile mosquitoes with beverton-holt survivability[J]. Mathematical Biosciences and Engineering, 2019, 16(2): 572-602. doi: 10.3934/mbe.2019028







 DownLoad:
DownLoad: