Citation: Sambasiva Rao Katuri, Rajesh Khanna. Kinetic growth model for hairy root cultures[J]. Mathematical Biosciences and Engineering, 2019, 16(2): 553-571. doi: 10.3934/mbe.2019027

| [1] | A. A. Dilorio, R. D. Cheetham and P. J. Weathers, Growth of transformed roots in a nutrient mist bioreactor: reactor performance and evaluation. Appl. Microbiol. Biot., 37 (1992), 457–462. |
| [2] | A. Chavarria-Krauser and U. Schurr, A cellular growth model for root tips. J. Theor. Biol., 230 (2004), 21–32. |
| [3] | B. H. Abbasi, R. Liu, P. K. Saxena and C. Z. Liu, Cichoric acid production from hairy root cultures of Echinacea purpurea grown in a modified airlift bioreactor. J. Chem. Tech. Biotechnol., 84 (2009), 1697–1701. |
| [4] | B. Han, J. C. Linden, N. P. Gujarathi and S. R. Wickramasinghe, Population balance approach to modeling hairy root growth. Biotechnol. Progr., 20 (2004), 872–879. |
| [5] | B. N. Mishra and R. Ranjan, Growth of hairy root cultures in various bioreactors for the production of secondary metabolites. Biotechnol. Appl. Bioc. 49 (2008), 1–10. |
| [6] | B. Neelwarne and S. B. Halagur, Red Beet: An Overview. In Red Beet Biotechnol., Springer US (2012), 1–43. |
| [7] | B. E. Wyslouzil, M. Whipple, C. Chatterjee, D. B. Walcerz, P. J. Weathers and D. P. Hart, Mist deposition onto hairy root cultures: aerosol modeling and experiments. Biotechnol. Progr., 13 (1997), 185–194. |
| [8] | C. Hackett and D. A. Rose, A model of the extension and branching of a seminal root of barley, and its use in studying relations between root dimensions I. The model. Aust. J. Biol. Sci., 25 (1972), 669–680. |
| [9] | C. Hackett and D. A. Rose, A model of the Extension and Branching of a Seminal Root of Barley, and Its Use in Studying Relations Between Root Dimensions II. Results and Inferences from Manipulation of the Model. Aust. J. Biol. Sci., 25 (1972), 681–690. |
| [10] | C. Z. Liu, Y. C. Wang, B. Zhao, C. Guo, F. Ouyang, H. C. Ye and G. F. Li, Development of a nutrient mist bioreactor for growth of hairy roots. In Vitro Cell. Dev-Pl. 35 (1999), 271–274. |
| [11] | D. Ramakrishnan and W. R. Curtis, Tricklebed root culture bioreactor design and scaleup: Growth, fluid dynamics, and oxygen mass transfer. Biotechnol. Bioeng., 88 (2004), 248–260. |
| [12] | J. Foreman and L. Dolan, Root hairs as a model system for studying plant cell growth. Ann. Bot., 88 (2001), 1–7. |
| [13] | K. C. Zygalakis, G. J. D. Kirk, D. L. Jones, M. Wissuwa and T. Roose, A dual porosity model of nutrient uptake by root hairs. New Phytol., 192 (2011), 676–688. |
| [14] | K. H. Kwok and P. M. Doran, Kinetic and stoichiometric analysis of hairy roots in a segmented bubble column reactor. Biotechnol. Progr. 11 (1995), 429–435. |
| [15] | L. S. Sung and S. Y. Huang, Lateral root bridging as a strategy to enhance LDOPA production in Stizolobium hassjoo hairy root cultures by using a mesh hindrance mist trickling bioreactor. Biotechnol. Bioeng., 94 (2006), 441–447. |
| [16] | M. Hjortso and U. Mukundan, Genetic transformation in Tagetes species (marigolds) for thiophene contents. Pl. Protoplasts Genet. Eng. V., Springer Berlin Heidelberg, (1994), 365–382. |
| [17] | M. A. Subroto and P. M. Doran, Production of steroidal alkaloidsby hairy roots of Solanum aviculare and the effect of gibberellic acid. Plant Cell Tiss. Org., 38 (1994), 93–102. |
| [18] | M. J. Towler, B. E. Wyslouzil and P. J. Weathers, Using an aerosol deposition model to increase hairy root growth in a mist reactor. Biotechnol. Bioeng., 96 (2007), 881–891. |
| [19] | P. Banyai, I. Blvnyos, L. Kursinszki and E. Szke, Cultivation of Lobelia inflate L. Hairy Root Culture in Bioreactor. In International Conference on Medicinal and Aromatic Plants (Part II), 597 (2001), 253–256. |
| [20] | P. Bastian, A. Chavarra-Krauser, C. Engwer, W. Jger, S. Marnach and M. Ptashnyk, Modelling in vitro growth of dense root networks. J. Theor Biol., 254 (2008), 99–109. |
| [21] | P. J. Weathers, C. Liu , M. Towler and B. E. Wyslouzil, Mist reactors: principles, comparison of various systems, and case studies. Electron. J. Integrative Biosciences, 3 (2008), 29–37. |
| [22] | P. T. Asplund and W. R. Curtis, Intrinsic oxygen use kinetics of transformed plant root culture. Biotechnol. Progr., 17 (2001), 481–489. |
| [23] | R. Ranjan, N. Ahmed, R. Khanna and B. N. Mishra, Design of an ON/OFF mist duty cycle in mist bioreactors for the growth of hairy roots. Biotechnol. Bioprocess Eng., 14 (2009), 38–45. |
| [24] | R. Ranjan, S. R. Katuri and R. Khanna, Discrete modeling of growth of hairy roots in a mist bioreactor. Chem. Eng. Technol., 38 (2015), 391–398. |
| [25] | S. Kim, E. Hopper and M. Hjortso, Hairy root growth models: effect of different branching patterns. Biotechnol. Progr., 11 (1995), 178–186. |
| [26] | S. H. Woo, J. M. Park and J. W. Yang, Induction of branch roots by cutting method in Hyoscyamus niger root culture. Plant Cell Tiss. Org., 48 (1997), 131–134. |
| [27] | S. Srivastava and A. K. Srivastava, Hairy root culture for mass-production of high-value secondary metabolites. Crit. Rev. Biotechnol., 27 (2007), 29–43. |
| [28] | S. Yu, and P. M. Doran, Oxygen requirements and mass transfer in hairyroot culture. Biotechnol. Bioeng., 44 (1994), 880–887. |
| [29] | T. L. Shiao and P. M. Doran, Root hairiness: effect on fluid flow and oxygen transfer in hairy root cultures. J. Biotechnol., 83 (2000), 199–210. |
| [30] | Y. J. Kim, P. J. Weathers and B. E. Wyslouzil, Growth dynamics of Artemisia annua hairy roots in three culture systems. Biotechnol. Bioeng., 83 (2003), 428–443. |
| [31] | Y. Kim, B. E. Wyslouzil and P. J. Weathers, Secondary metabolism of hairy root cultures in bioreactors. In Vitro Cell. Dev-Pl., 38 (2002), 1–10. |
| [32] | Y. Kim, B. E. Wyslouzil and P. J. Weathers, A comparative study of mist and bubble column reactors in the in vitro production of Artemisinin. Plant Cell Rep., 20 (2001), 451–455. |
| [33] | Y. Nakashimada, U. Nobuyuki, and K. Takeshi, Stimulation of emergence of root apical meristems in horseradish hairy root by auxin supplementation and its kinetic model. J. Ferment. Bioeng., 77 (1994), 178–182. |
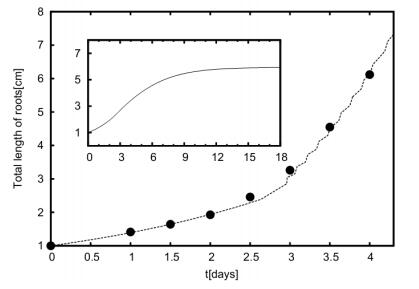
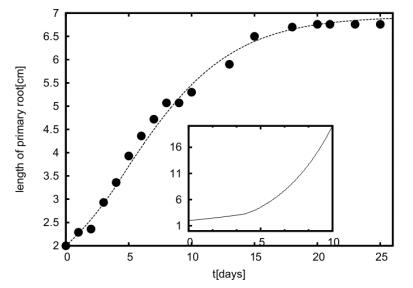
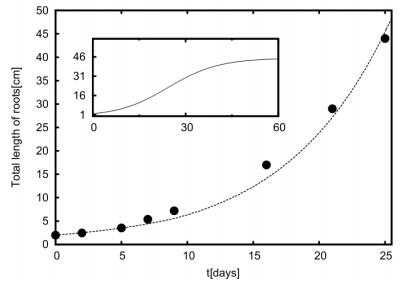
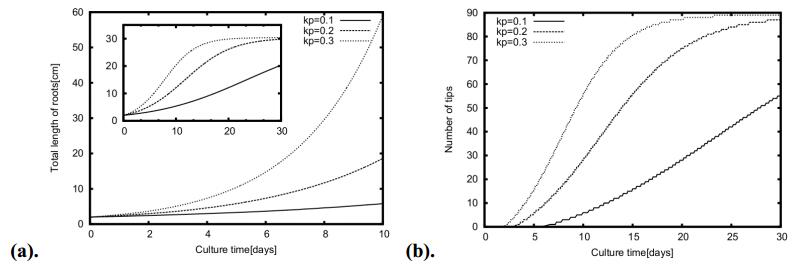
Figures(11) / Tables(1)

Sambasiva Rao Katuri, Rajesh Khanna. Kinetic growth model for hairy root cultures[J]. Mathematical Biosciences and Engineering, 2019, 16(2): 553-571. doi: 10.3934/mbe.2019027







 DownLoad:
DownLoad: