Citation: Jan Willem Erisman, Nick van Eekeren, Jan de Wit, Chris Koopmans, Willemijn Cuijpers, Natasja Oerlemans, Ben J. Koks. Agriculture and biodiversity: a better balance benefits both[J]. AIMS Agriculture and Food, 2016, 1(2): 157-174. doi: 10.3934/agrfood.2016.2.157

| [1] | Convention on Biological Diversity, United Nations. 1992. Available from: https://www.cbd.int/doc/legal/cbd-en.pdf |
| [2] |
Fahrig L, Baudry J, Brotons L, et al. (2011) Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol Lett 14: 101-112. doi: 10.1111/j.1461-0248.2010.01559.x

|
| [3] |
Odum EP (1969) The strategy of ecosystem development. Science 164: 262-270 doi: 10.1126/science.164.3877.262
|
| [4] |
Holling CS (1973) Resilience and stability of ecological systems. Annu Rev Ecol Syst 4: 1-23. doi: 10.1146/annurev.es.04.110173.000245
|
| [5] |
Beintema AJ (1986) Man-made Polders in the Netherlands: a Traditional Habitat for shorebirds. Colon Waterbirds 9: 196-202 doi: 10.2307/1521213
|
| [6] | EEA (2015) European ecosystem assessment — concept, data, and implementation. Contribution to Target 2 Action 5 Mapping and Assessment of Ecosystems and their Services (MAES) of the EU Biodiversity Strategy to 2020. EEA Technical Report no.6/2015, EEA, Copenhagen, Denmark. |
| [7] |
Henckel L, Borger L, Meiss H, et al. (2015) Organic fields sustain weed metacommunity dynamics in farmland landscapes. Proc R Soc B 282: 20150002. doi: 10.1098/rspb.2015.0002
|
| [8] | Wereld Natuur Fonds (2015) Living Planet Report. Natuur in Nederland. WNF, Zeist, The Netherlands. |
| [9] | Buckwell A, Uhre AN, Williams A, et al. (2012) Sustainable Intensification of European Agriculture A review sponsored by the RISE Foundation. www.risefoundation.eu |
| [10] |
Geiger F, Bengtsson J, Berendse F, et al. (2010) Persistent negative effects of pesticides and biological control potential on European farmland. Basic Appl Ecol 11: 97-105. doi: 10.1016/j.baae.2009.12.001
|
| [11] |
Foley JA, DeFries R, Asner GP, et al. (2005) Global consequences of land use. Science 309: 570-574. doi: 10.1126/science.1111772
|
| [12] |
Lechenet M, Bretagnolle V, Bockstaller C, et al. (2014) Reconciling pesticide reduction with economic and environmental sustainability in arable farming. PLoS ONE 9: e97922. doi: 10.1371/journal.pone.0097922
|
| [13] | FAO, WFP and IFAD (2012) The State of Food Insecurity in the World 2012. Economic growth is necessary but not sufficient to accelerate reduction of hunger and malnutrition. Rome, FAO. |
| [14] | Tittonell P (2013) Farming Systems Ecology. Towards ecological intensification of world agriculture. Inaugural lecture upon taking up the position of Chair in Farming Systems Ecology at Wageningen University on 16 May 2013. http://www.wageningenur.nl/en/show/Feeding-the-world-population-sustainably-and- efficiently-with-ecologically-intensive-agriculture.htm. |
| [15] |
Tomich TP, Brodt S, Ferris H, et al. (2011) Agroecology: A Review from a Global-Change Perspective. Annu Rev Environ Resour 36: 193-222. doi: 10.1146/annurev-environ-012110-121302
|
| [16] |
Davis AS, Hill JD, Chase CA, et al. (2012) Increasing Cropping System Diversity Balances Productivity, Profitability and Environmental Health. PLoS ONE 7: e47149. doi: 10.1371/journal.pone.0047149
|
| [17] |
Tilman D, Wedin D, Knops J (1996) Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature 379: 718-720. doi: 10.1038/379718a0
|
| [18] | Tilman D, Reich P, Isbell F (2012) Biodiversity impacts ecosystem productivity as much as resources, disturbance or herbivory. Proc Natl Acad Sci 109: 10394-10397. |
| [19] | FAO (2010) Sustainable Diets and Biodiversity. Directions and Solutions for Policy, Research and Action. Proceedings of the International Scientific Symposium ‘Biodiversity and Sustainable Diets United Against Hunger’ on 3-5 November 2010 Food and Agriculture Organization of the United Nations, Rome. |
| [20] |
Cardinale BJ, Duffy JE, Gonzalez A, et al. (2012) Biodiversity loss and its impact on humanity. Nature 486: 59-67. doi: 10.1038/nature11148
|
| [21] | Fischer J, Lindenmayer DB, Manning AD (2006) Biodiversity, ecosystem function, and resilience: ten guiding principles for commodity production landscapes. Front Ecol Environ 4: 80-86. |
| [22] | Huyghe C, Litrico I, Surault F (2012) Agronomic value and provisioning services of multi-species swards. Grassl Sci Eur 17: 35-46. |
| [23] | Schmid B (2002) The species richness-productivity controversy. Trends Ecol Evol 17: 113-114. |
| [24] |
Zuppinger-Dingley D, Schmid B, Petermann JS, et al. (2014) Selection for niche differentiation in plant communities increases biodiversity effects. Nature 515: 108-111. doi: 10.1038/nature13869
|
| [25] |
Vandermeer J, van Noordwijk M, Anderson J, et al. (1998) Global change and multi-species agroecosystems: Concepts and issues. Agric Ecosyst Environ 67: 1-22. doi: 10.1016/S0167-8809(97)00150-3
|
| [26] |
Walker B (1995) Conserving biological diversity through ecosystem resilience. Conserv Biol 9: 747-752. doi: 10.1046/j.1523-1739.1995.09040747.x
|
| [27] | Jacobsen SE, Sørensen M, Pedersen S, et al. (2013) Feeding the world: genetically modified crops versus agricultural biodiversity. Agron Sustain Dev 33: 651-662. |
| [28] | Altieri MA (2009) Agroecology, small farms and food sovereignty. Mon Rev 61: 102. |
| [29] |
Mundt CC (2002) Use of multiline cultivars and cultivar mixtures for disease management. Annu Rev Phytopathol 40: 381-410. doi: 10.1146/annurev.phyto.40.011402.113723
|
| [30] |
Wolfe MS (2000) Crop strength through diversity. Nature 406: 681-682. doi: 10.1038/35021152
|
| [31] | Jareño D, Viñuela J, Luque-Larena JJ, et al. (2015) Factors associated with the colonization of agricultural areas by common voles Microtus arvalis in NW Spain. Biol Invasions 17: 2315-2327. |
| [32] | Evans KL (2004) The potential for interactions between predation and habitat change to cause population declines of farmland birds. Ibis 146: 1-13. |
| [33] |
Kentie R, Both C, Hooijmeijer JCE, et al. (2015) Management of modern agricultural landscapes increases nest predation rates in Black-tailed Godwits Limosa limosa. Ibis 157: 614-625. doi: 10.1111/ibi.12273
|
| [34] | Bretagnolle V, Gauffre B, Meiss H, et al. (2011) The Role of Grassland Areas within Arable Cropping Systems for the Conservation of Biodiversity at the Regional Level. In: Grassland Productivity and Ecosystem Services, ed. Lemaire, G. et al. 251-60. USA: CABI |
| [35] |
Pywell RF, Heard MS, Woodcock BA, et al. (2015) Wildlife friendly farming increases crop yield: evidence for ecological intensification. Proc R Soc B 282: 20151740. doi: 10.1098/rspb.2015.1740
|
| [36] | Compendium, 2015. Available from: https://www.Compendiumvoordeleefomgeving.nl |
| [37] |
Donald PF, Sanderson FJ, Burfield IJ, et al. (2006) Further evidence of continent-wide impacts of agricultural intensification on European farmland birds, 1990-2000. Agric Ecosyst Environ 116: 189-196. doi: 10.1016/j.agee.2006.02.007
|
| [38] | Ministry of Economic Affairs (2014) Convention on Biological Diversity. Fifth National Report of the Kingdom of the Netherlands. Den Haag, The Netherlands. |
| [39] | PBL (2014) Balans van de Leefomgeving 2014. De toekomst is nú, Den Haag: Planbureau voor de Leefomgeving. The Netherlands. |
| [40] | Dise NB, Ashmore M, Belyazid S, et al. (2011) Nitrogen as a threat to European terrestrial biodiversity. In: The European Nitrogen Assessment, ed. Sutton MA, Howard CM, Erisman JW, et al. Cambridge University Press. |
| [41] | Erisman JW, Galloway JN, Dise NB, et al. (2015) Nitrogen: Too much of a vital resource. WWF Netherlands, Zeist, the Netherlands. 27. |
| [42] |
Van den Noort PC (1987) Land consolidation in the Netherlands. Land Use Policy 4: 11-13. doi: 10.1016/0264-8377(87)90004-4
|
| [43] | Napel J, Bianchi F, Bestman MWP (2006) Utilising intrinsic robustness in agricultural production systems. TransForum Working Papers. 32-53. (available at www.louisbolk.org) |
| [44] |
Erisman JW, Brasseur G, Ciais P, et al. (2015) Global change: Put people at the centre of global risk management. Nature 519: 151-153. doi: 10.1038/519151a
|
| [45] | Altieri MA (1999) The ecological role of biodiversity in agroecosystems. Agric Ecosyst Environ 74: 19-31. |
| [46] | Rodale Institute (2011) The farming systems trial. Celebrating 30 years. The Rodale Institute, USA. Available from: http://rodaleinstitute.org/assets/FSTbooklet.pdf |
| [47] | De Schutter O (2010) Report submitted by the Special Rapporteur on the right to food, Olivier De Schutter. United Nations, General Assembly. Human Rights Council, Sixteenth session, Agenda item 3: Promotion and protection of all human rights, civil, political, economic, social and cultural rights, including the right to development. Available from: www2.ohchr.org/english/issues/food/docs/A-HRC-16-49. pdf). August 17, 2011. |
| [48] | Deru J, Schilder H, Van der Schoot JR, et al. (2014) Genetic differences in root mass of Lolium perenne varieties under eld conditions. Euphytica 199: 223-232. |
| [49] |
Gewin V (2010) An underground revolution. Nature 466: 552-553. doi: 10.1038/466552a
|
| [50] |
Van Grinsven HJM, Holland M, Jacobsen BH, et al. (2013) Costs and Benefits of Nitrogen for Europe and Implications for Mitigation. Environ Sci Technol 47: 3571-3579. doi: 10.1021/es303804g
|
| [51] | Lin BB (2011) Resilience in Agriculture through Crop Diversification: Adaptive Management for Environmental Change. BioScience 61: 183-193. |
| [52] |
De Ponti T, Rijk B, Van Ittersum MK (2012) The crop yield gap between organic and conventional agriculture. Agric Syst 108: 1-9. doi: 10.1016/j.agsy.2011.12.004
|
| [53] |
Seufert V, Ramankutty N, Foley JA (2012) Comparing the yields of organic and conventional agriculture. Nature 485: 229-232 doi: 10.1038/nature11069
|
| [54] | Ponisio LC, M’Gonigle LK, Mace KC, et al. (2015) Diversification practices reduce organic to conventional yield gap. Proc R Soc B 282: 20141396. |
| [55] |
Altieri MA, Nicholls CI, Henao A, et al. (2015) Agroecology and the design of climate change-resilient farming systems. Agron Sustain Dev 35: 869-890 doi: 10.1007/s13593-015-0285-2
|
| [56] |
Isbell F, Craven D, Connolly J, et al. (2015) Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature 526: 574-577. doi: 10.1038/nature15374
|
| [57] | Nemes N (2009) Comparative analysis of organic and non-organic farming systems: a critical assessment of farm profitability. FAO, Rome, Italy. |
| [58] |
Gonthier DJ, Ennis KJ, Farinas S, et al. (2014) Biodiversity conservation in agriculture requires a multi-scale approach. Proc R Soc B 281: 20141358. doi: 10.1098/rspb.2014.1358
|
| [59] |
Smith HG, Dänhardt J, Lindström A, et al. (2010) Consequences of organic farming and landscape heterogeneity for species richness and abundance of farmland birds. Oecologia 162: 1071-1079 doi: 10.1007/s00442-010-1588-2
|
| [60] | Smits MJW, van Alebeek FAN (2007) Biodiversiteit en kleine landschapselementen in de biologische landbouw; Een literatuurstudie. Wageningen, Wettelijke Onderzoekstaken Natuur & Milieu, WOT-rapport 39. 84. |
| [61] | Schneider MK, Lüscher G, Jeanneret P, et al. (2014) Gains to species diversity in organically farmed fields are not propagated at the farm level. Nat Commun 5: 1-9. |
| [62] | Kretschmer H, Pfeffer H, Hoffmann J, et al. (1995) Strukturelemente in Agrarlandschaften Ostdeutschlands - Bedeutung für den Biotop- und Artenschutz. ZALF-Bericht 19, Selbstverlag ZALF, Müncheberg: 164 S. |
| [63] | Hoffmann J, Kretschmer H, Pfeffer H (1999) Effects of Current Landscape Patterns on Biodiversity. -In: Tenhunen, J.D., R. Lenz und R.E. Hantschel (Hrsg.): Ecosystem properties and landscape function in Central Europe. Ecological Studies. Springer. |
| [64] |
Benton TG, Vickery JA, Wilson JD (2003) Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol Evol 18: 182-188. doi: 10.1016/S0169-5347(03)00011-9
|
| [65] |
Allan E, Bossdorf O, Dormann CF, et al. (2014) Interannual variation in land-use intensity enhances grassland multidiversity. Proc Natl Acad Sci 111: 308-313. doi: 10.1073/pnas.1312213111
|
| [66] | Payne RJ, Thompson AM, Standen V, et al. (2012) Impact of simulated nitrogen pollution on heathland microfauna, mesofauna and plants. Eur J Soil Biol 49: 73-79. |
| [67] | Wymenga E, Latour J, Beemster N, et al. (2015) Terugkerende muizenplagen in Nederland. Inventarisatie, sturende factoren en beheersing. A&W-rapport 2123. Altenburg & Wymenga bv, Alterra Wageningen UR, Livestock Research Wageningen, Wetterskip Fryslân, Stichting Werkgroep Grauwe Kiekendief. Feanwâlden. |
| [68] | Van Eekeren N, Heeres E, Smeding F (2003) Leven onder de graszode - Discussiestuk over het beoordelen en beïnvloeden van bodemleven in de biologische melkveehouderij. Louis Bolk publikatie LV 52. Louis Bolk Instituut Driebergen, The Netherlands. |
| [69] |
Van Groenigen JW, Lubbers IM, Vos HMJ, et al. (2014) Earthworms increase plant production: a meta-analysis. Sci Reports 4: 6365. doi: 10.1038/srep06365
|
| [70] | Geiger F (2011) Agricultural intensification and farmland birds. Thesis Wageningen University, The Netherlands. |
| [71] | Brink C, van Grinsven H, Jacobsen BH, et al. (2011) Costs and benefits of nitrogen in the environment. In: Sutton MA, Howard CM, Erisman JW, et al. (Eds). The European Nitrogen Assessment. Cambridge, UK, Cambridge University Press, Chapter 19, pp. 434-462. |
| [72] |
Compton JE, Harrison JA, Dennis RL, et al. (2011) Ecosystem services altered by human changes in the nitrogen cycle: a new perspective for US decision making. Ecol Lett 14: 804-815. doi: 10.1111/j.1461-0248.2011.01631.x
|
| [73] |
Frison EA, Cherfas J, Hodgkin T (2011) Agricultural biodiversity is essential for a sustainable improvement in food and nutrition security. Sustainability 3: 238-253. doi: 10.3390/su3010238
|
| [74] |
Lemaire G, Franzluebbers A, Carvalho PCF, et al. (2014) Integrated crop-livestock systems: Strategies to achieve synergy between agricultural production and environmental quality. Agric Ecosyst Environ 190: 4-8. doi: 10.1016/j.agee.2013.08.009
|
| [75] |
Grace JB, Anderson TM, Seabloom EW, et al. (2016) Integrative modelling reveals mechanisms linking productivity and plant species richness. Nature 529: 390-393. doi: 10.1038/nature16524
|
| [76] | Van Eekeren N, Verhoeven F, Erisman JW (2015) Verkenning Kritische Prestatie Indicatoren voor stimulering van een biodiverse melkveehouderij. Louis Bolk, Driebergen, The Netherlands. |
| [77] |
Groot JCJ, Rossing WAH, Jellema A, et al. (2007) Exploring multi-scale trade-offs between nature conservation, agricultural profits and landscape quality - a methodology to support discussions on land-use perspectives. Agric Ecosyst Environ 120: 58-69. doi: 10.1016/j.agee.2006.03.037
|
| [78] | De Snoo GR, Herzon I, Staats H, et al. (2012) Toward effective nature conservation on farmland: making farmers matter. Conserv Lett 6: 66-72. |
| [79] | Duru M, Therond O, Martin G (2015) How to implement biodiversity-based agriculture to enhance ecosystem services: A review. Agron Sustain Dev 35: 1259-1281. |

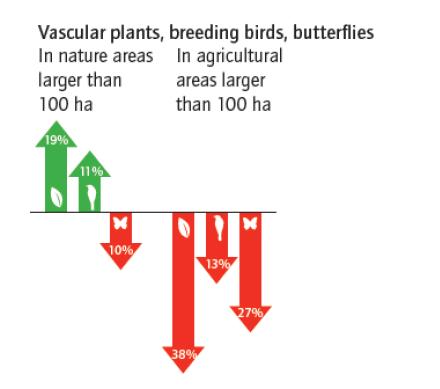
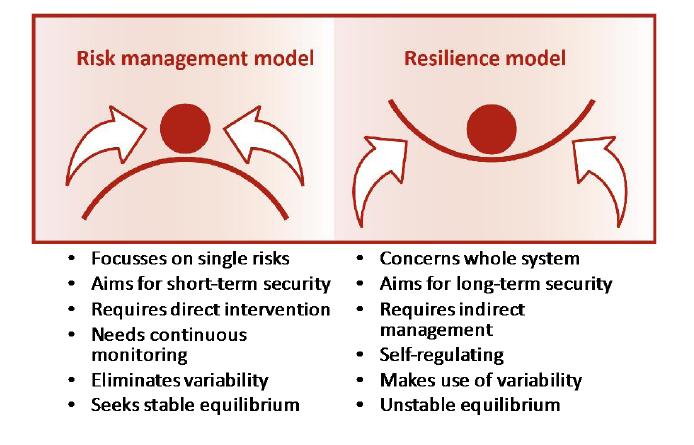
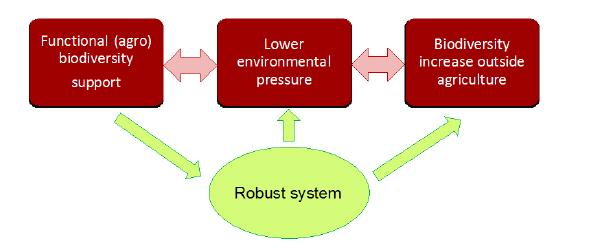
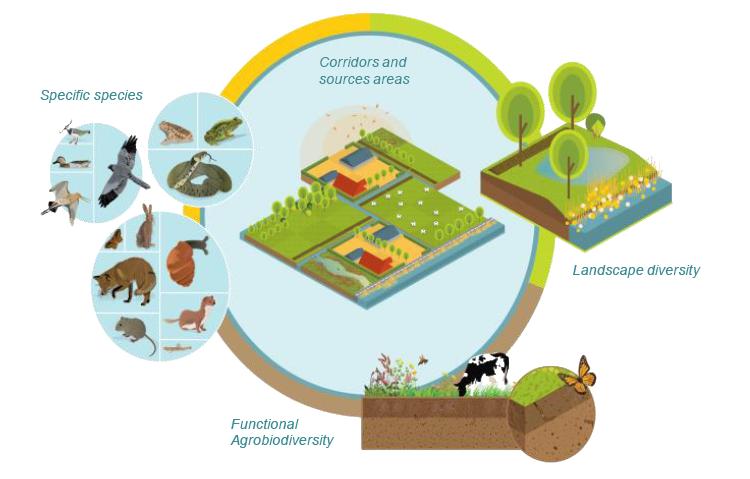
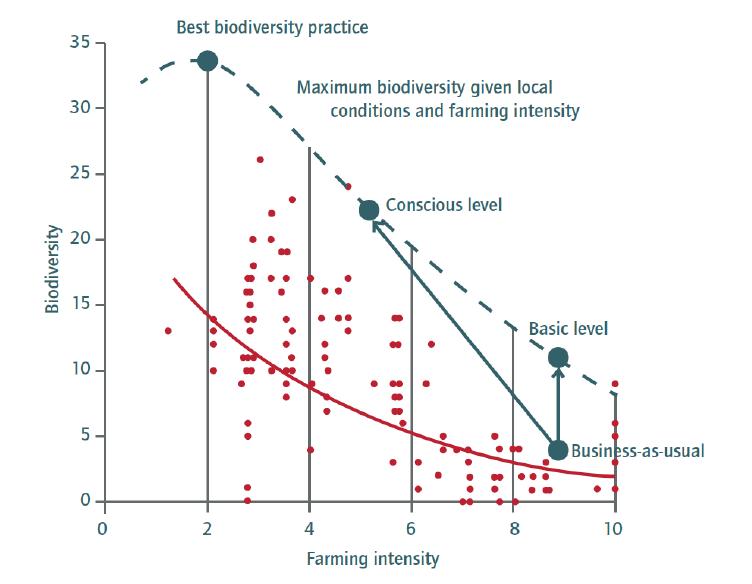
Figures(6)

Jan Willem Erisman, Nick van Eekeren, Jan de Wit, Chris Koopmans, Willemijn Cuijpers, Natasja Oerlemans, Ben J. Koks. Agriculture and biodiversity: a better balance benefits both[J]. AIMS Agriculture and Food, 2016, 1(2): 157-174. doi: 10.3934/agrfood.2016.2.157







 DownLoad:
DownLoad: