Citation: Liwei Chen, Jaslyn Lee, Wei Ning Chen. The use of metabolic engineering to produce fatty acid-derived biofuel and chemicals in Saccharomyces cerevisiae: a review[J]. AIMS Bioengineering, 2016, 3(4): 468-492. doi: 10.3934/bioeng.2016.4.468

| [1] |
Handke P, Lynch SA, Gill RT (2011) Application and engineering of fatty acid biosynthesis in Escherichia coli for advanced fuels and chemicals. Metab Eng 13: 28–37. doi: 10.1016/j.ymben.2010.10.007

|
| [2] |
Pfleger BF, Gossing M, Nielsen J (2015) Metabolic engineering strategies for microbial synthesis of oleochemicals. Metab Eng 29: 1–11. doi: 10.1016/j.ymben.2015.01.009
|
| [3] | Zhou YJ, Buijs NA, Siewers V, et al. (2014) Fatty Acid-Derived Biofuels and Chemicals Production in Saccharomyces cerevisiae. Front Bioeng Biotechnol 2: 32. |
| [4] |
Lennen RM, Pfleger BF (2013) Microbial production of fatty acid-derived fuels and chemicals. Curr Opin Biotechnol 24: 1044–1053. doi: 10.1016/j.copbio.2013.02.028
|
| [5] |
Torella JP, Ford TJ, Kim SN, et al. (2013) Tailored fatty acid synthesis via dynamic control of fatty acid elongation. Proc Natl Acad Sci USA 110: 11290–11295. doi: 10.1073/pnas.1307129110
|
| [6] | Howard TP, Middelhaufe S, Moore K, et al. (2013) Synthesis of customized petroleum-replica fuel molecules by targeted modification of free fatty acid pools in Escherichia coli. Proc Natl Acad Sci USA 110: 7636–7641. |
| [7] |
Choi YJ, Lee SY (2013) Microbial production of short-chain alkanes. Nature 502: 571–574. doi: 10.1038/nature12536
|
| [8] | Li M, Borodina I (2015) Application of synthetic biology for production of chemicals in yeast Saccharomyces cerevisiae. FEMS Yeast Res 15: 3281–3285. |
| [9] |
Cherry JM, Hong EL, Amundsen C, et al. (2012) Saccharomyces Genome Database: the genomics resource of budding yeast. Nucleic Acids Res 40: D700–D705. doi: 10.1093/nar/gkr1029
|
| [10] |
Krivoruchko A, Siewers V, Nielsen J (2011) Opportunities for yeast metabolic engineering: Lessons from synthetic biology. Biotechnol J 6: 262–276. doi: 10.1002/biot.201000308
|
| [11] | Blazeck J, Hill A, Liu L, et al. (2014) Harnessing Yarrowia lipolytica lipogenesis to create a platform for lipid and biofuel production. Nat Commun 5: 3131. |
| [12] |
Lin X, Wang Y, Zhang S, et al. (2014) Functional integration of multiple genes into the genome of the oleaginous yeast Rhodosporidium toruloides. FEMS Yeast Res 14: 547–555. doi: 10.1111/1567-1364.12140
|
| [13] |
Liu Y, Koh CMJ, Ngoh ST, et al. (2015) Engineering an efficient and tight D-amino acid-inducible gene expression system in Rhodosporidium/Rhodotorula species. Microb Cell Fact 14: 1–16. doi: 10.1186/s12934-014-0183-3
|
| [14] |
Feng X, Lian J, Zhao H (2015) Metabolic engineering of Saccharomyces cerevisiae to improve 1-hexadecanol production. Metab Eng 27: 10–19. doi: 10.1016/j.ymben.2014.10.001
|
| [15] | Zhou YJ, Buijs NA, Zhu Z, et al. (2016) Production of fatty acid-derived oleochemicals and biofuels by synthetic yeast cell factories. Nat Commun 7. |
| [16] |
Nozzi NE, Desai SH, Case AE, et al. (2014) Metabolic engineering for higher alcohol production. Metab Eng 25: 174–182. doi: 10.1016/j.ymben.2014.07.007
|
| [17] | Kang MK, Nielsen J (2016) Biobased production of alkanes and alkenes through metabolic engineering of microorganisms. J Ind Microbiol Biotechnol: 1–10. |
| [18] | McNaught AD, Wilkinson A (1997) IUPAC Compendium of Chemical Terminology, 2Eds. |
| [19] |
Tehlivets O, Scheuringer K, Kohlwein SD (2007) Fatty acid synthesis and elongation in yeast. Biochim Biophys Acta Mol Cell Biol Lipids 1771: 255–270. doi: 10.1016/j.bbalip.2006.07.004
|
| [20] |
Wakil SJ, Stoops JK, Joshi VC (1983) Fatty Acid Synthesis and its Regulation. Annu Rev Biochem 52: 537–579. doi: 10.1146/annurev.bi.52.070183.002541
|
| [21] |
Al-Feel W, Chirala SS, Wakil SJ (1992) Cloning of the yeast FAS3 gene and primary structure of yeast acetyl-CoA carboxylase. Proc Natl Acad Sci USA 89: 4534–4538. doi: 10.1073/pnas.89.10.4534
|
| [22] | Woods A, Munday MR, Scott J, et al. (1994) Yeast SNF1 is functionally related to mammalian AMP-activated protein kinase and regulates acetyl-CoA carboxylase in vivo. J Biol Chem 269: 19509–19515. |
| [23] |
Jenni S, Leibundgut M, Maier T, et al. (2006) Architecture of a Fungal Fatty Acid Synthase at 5 A Resolution. Science 311: 1263–1267. doi: 10.1126/science.1123251
|
| [24] |
Leibundgut M, Maier T, Jenni S, et al. (2008) The multienzyme architecture of eukaryotic fatty acid synthases. Curr Opin Struct Biol 18: 714–725. doi: 10.1016/j.sbi.2008.09.008
|
| [25] |
Lomakin IB, Xiong Y, Steitz TA (2007) The crystal structure of yeast fatty acid synthase, a cellular machine with eight active sites working together. Cell 129: 319–332. doi: 10.1016/j.cell.2007.03.013
|
| [26] |
Leibundgut M, Jenni S, Frick C, et al. (2007) Structural basis for substrate delivery by acyl carrier protein in the yeast fatty acid synthase. Science 316: 288–290. doi: 10.1126/science.1138249
|
| [27] |
Jenni S, Leibundgut M, Boehringer D, et al. (2007) Structure of Fungal Fatty Acid Synthase and Implications for Iterative Substrate Shuttling. Science 316: 254–261. doi: 10.1126/science.1138248
|
| [28] |
Gipson P, Mills DJ, Wouts R, et al. (2010) Direct structural insight into the substrate-shuttling mechanism of yeast fatty acid synthase by electron cryomicroscopy. Proc Natl Acad Sci USA 107: 9164–9169. doi: 10.1073/pnas.0913547107
|
| [29] | Magnuson K, Jackowski S, Rock CO, et al. (1993) Regulation of fatty acid biosynthesis in Escherichia coli. Microbiol Rev 57: 522–542. |
| [30] |
Brignole EJ, Smith S, Asturias FJ (2009) Conformational flexibility of metazoan fatty acid synthase enables catalysis. Nat Struct Mol Biol 16: 190–197. doi: 10.1038/nsmb.1532
|
| [31] |
Oh CS, Toke DA, Mandala S, et al. (1997) ELO2 and ELO3, homologues of the Saccharomyces cerevisiae ELO1 gene, function in fatty acid elongation and are required for sphingolipid formation. J Biol Chem 272: 17376–17384. doi: 10.1074/jbc.272.28.17376
|
| [32] |
Dittrich F, Zajonc D, Hühne K, et al. (1998) Fatty acid elongation in yeast. Eur J Biochem 252: 477–485. doi: 10.1046/j.1432-1327.1998.2520477.x
|
| [33] |
Santiago TC, Mamoun CB (2003) Genome Expression Analysis in Yeast Reveals Novel Transcriptional Regulation by Inositol and Choline and New Regulatory Functions for Opi1p, Ino2p, and Ino4p. J Biol Chem 278: 38723–38730. doi: 10.1074/jbc.M303008200
|
| [34] |
Loewen CJR, Gaspar ML, Jesch SA, et al. (2004) Phospholipid Metabolism Regulated by a Transcription Factor Sensing Phosphatidic Acid. Science 304: 1644–1647. doi: 10.1126/science.1096083
|
| [35] |
Chirala SS, Zhong Q, Huang W, et al. (1994) Analysis of FAS3/ACC regulatory region of Saccharomyces cerevisiae: identification of a functional UASINO and sequences responsible for fatty acid mediated repression. Nucleic Acids Res 22: 412–418. doi: 10.1093/nar/22.3.412
|
| [36] |
Chirala SS (1992) Coordinated regulation and inositol-mediated and fatty acid-mediated repression of fatty acid synthase genes in Saccharomyces cerevisiae. Proc Natl Acad Sci USA 89: 10232–10236. doi: 10.1073/pnas.89.21.10232
|
| [37] |
Jesch SA, Liu P, Zhao X, et al. (2006) Multiple Endoplasmic Reticulum-to-Nucleus Signaling Pathways Coordinate Phospholipid Metabolism with Gene Expression by Distinct Mechanisms. J Biol Chem 281: 24070–24083. doi: 10.1074/jbc.M604541200
|
| [38] |
Jesch SA, Zhao X, Wells MT, et al. (2005) Genome-wide analysis reveals inositol, not choline, as the major effector of Ino2p-Ino4p and unfolded protein response target gene expression in yeast. J Biol Chem 280: 9106–9118. doi: 10.1074/jbc.M411770200
|
| [39] |
Huisinga KL, Pugh BF (2004) A genome-wide housekeeping role for TFIID and a highly regulated stress-related role for SAGA in Saccharomyces cerevisiae. Mol Cell 13: 573–585. doi: 10.1016/S1097-2765(04)00087-5
|
| [40] | Greenberg ML, Lopes JM (1996) Genetic regulation of phospholipid biosynthesis in Saccharomyces cerevisiae. Microbiol Rev 60: 1–20. |
| [41] |
Shirra MK, Pattonvogt J, Ulrich A, et al. (2001) Inhibition of acetyl coenzyme a carboxylase activity restores expression of the INO1 gene in a snf1 mutant strain of Saccharomyces cerevisiae. Mol Cell Biol 21: 5710–5722. doi: 10.1128/MCB.21.17.5710-5722.2001
|
| [42] |
Wenz P, Schwank S, Hoja U, et al. (2001) A downstream regulatory element located within the coding sequence mediates autoregulated expression of the yeast fatty acid synthase gene FAS2 by the FAS1 gene product. Nucleic Acids Res 29: 4625–4632. doi: 10.1093/nar/29.22.4625
|
| [43] |
Fichtlscherer F, Wellein C, Mittag M, et al. (2000) A novel function of yeast fatty acid synthase. Eur J Biochem 267: 2666–2671. doi: 10.1046/j.1432-1327.2000.01282.x
|
| [44] |
Faergeman NJ, Knudsen J (1997) Role of long-chain fatty acyl-CoA esters in the regulation of metabolism and in cell signalling. Biochem J 323: 1–12. doi: 10.1042/bj3230001
|
| [45] |
Hedbacker K, Carlson M (2008) SNF1/AMPK pathways in yeast. Front Biosci 13: 2408–2420. doi: 10.2741/2854
|
| [46] |
Griffin TJ, Gygi SP, Ideker T, et al. (2002) Complementary profiling of gene expression at the transcriptome and proteome levels in Saccharomyces cerevisiae. Mol Cell Proteomics 1: 323–333. doi: 10.1074/mcp.M200001-MCP200
|
| [47] | Martin CE, Oh CS, Jiang Y (2007) Regulation of long chain unsaturated fatty acid synthesis in yeast. BBA-Mol Cell Biol L 1771: 271–285. |
| [48] |
DeRisi JL, Iyer VR, Brown PO (1997) Exploring the metabolic and genetic control of gene expression on a genomic scale. Science 278: 680–686. doi: 10.1126/science.278.5338.680
|
| [49] |
Nakagawa Y, Sakumoto N, Kaneko Y, et al. (2002) Mga2p is a putative sensor for low temperature and oxygen to induce OLE1 transcription in Saccharomyces cerevisiae. Biochem Biophys Res Commun 291: 707–713. doi: 10.1006/bbrc.2002.6507
|
| [50] |
Choi JY, Stukey J, Hwang SY, et al. (1996) Regulatory elements that control transcription activation and unsaturated fatty acid-mediated repression of the Saccharomyces cerevisiae OLE1 gene. J Biol Chem 271: 3581–3589. doi: 10.1074/jbc.271.7.3581
|
| [51] |
Gonzalez CI, Martin CE (1996) Fatty acid-responsive control of mRNA stability: unsaturated fatty acid-induced degradation of the Saccharomyces OLE1 transcript. J Biol Chem 271: 25801–25809. doi: 10.1074/jbc.271.42.25801
|
| [52] | Vasconcelles MJ, Jiang Y, McDaid K, et al. (2001) Identification and characterization of a low oxygen response element (LORE) involved in the hypoxic induction of a family of S.cerevisiae genes: implications for the conservation of oxygen sensing in eukaryotes. J Biol Chem 276: 14374–14384. |
| [53] |
Puig S, Askeland E, Thiele DJ (2005) Coordinated remodeling of cellular metabolism during iron deficiency through targeted mRNA degradation. Cell 120: 99–110. doi: 10.1016/j.cell.2004.11.032
|
| [54] |
Beopoulos A, Nicaud JM, Gaillardin C (2011) An overview of lipid metabolism in yeasts and its impact on biotechnological processes. Appl Microbiol Biotechnol 90: 1193–1206. doi: 10.1007/s00253-011-3212-8
|
| [55] | Black PN, DiRusso CC (2007) Yeast acyl-CoA synthetases at the crossroads of fatty acid metabolism and regulation. BBA-Mol Cell Biol L 1771: 286–298. |
| [56] |
Hiltunen JK, Mursula AM, Rottensteiner H, et al. (2003) The biochemistry of peroxisomal beta-oxidation in the yeast Saccharomyces cerevisiae. FEMS Microbiol Rev 27: 35–64. doi: 10.1016/S0168-6445(03)00017-2
|
| [57] |
van Roermund CWT, Waterham HR, Ijlst L, et al. (2003) Fatty acid metabolism in Saccharomyces cerevisiae. Cell Mol Life Sci 60: 1838–1851. doi: 10.1007/s00018-003-3076-x
|
| [58] | Hettema EH, van Roermund CW, Distel B, et al. (1996) The ABC transporter proteins Pat1 and Pat2 are required for import of long-chain fatty acids into peroxisomes of Saccharomyces cerevisiae. EMBO J 15: 3813–3822. |
| [59] |
Dmochowska A, Dignard D, Maleszka R, et al. (1990) Structure and transcriptional control of the Saccharomyces cerevisiae POX1 gene encoding acylcoenzyme A oxidase. Gene 88: 247–252. doi: 10.1016/0378-1119(90)90038-S
|
| [60] | Wang HJ, Le Dall MT, Waché Y, et al. (1999) Evaluation of acyl coenzyme A oxidase (Aox) isozyme function in the n- alkane-assimilating yeast Yarrowia lipolytica. J Bacteriol 181: 5140–5148. |
| [61] |
Mlickova K, Roux E, Athenstaedt K, et al. (2004) Lipid accumulation, lipid body formation, and acyl coenzyme A oxidases of the yeast Yarrowia lipolytica. Appl Environ Microbiol 70: 3918–3924. doi: 10.1128/AEM.70.7.3918-3924.2004
|
| [62] |
Mlickova K, Luo Y, d'Andrea S, et al. (2004) Acyl-CoA oxidase, a key step for lipid accumulation in the yeast Yarrowia lipolytica. J Mol Catal B -Enzym 28: 81–85. doi: 10.1016/j.molcatb.2004.01.007
|
| [63] |
Lennen RM, Braden DJ, West RM, et al. (2010) A process for microbial hydrocarbon synthesis: overproduction of fatty acids in Escherichia coli and catalytic conversion to alkanes. Biotechnol Bioeng 106: 193–202. doi: 10.1002/bit.22660
|
| [64] |
Liu T, Vora H, Khosla C (2010) Quantitative analysis and engineering of fatty acid biosynthesis in E. coli. Metab Eng 12: 378–386. doi: 10.1016/j.ymben.2010.02.003
|
| [65] |
Liu H, Yu C, Feng D, et al. (2012) Production of extracellular fatty acid using engineered Escherichia coli. Microb Cell Fact 11: 1–13. doi: 10.1186/1475-2859-11-1
|
| [66] |
Lennen RM, Pfleger BF (2012) Engineering Escherichia coli to synthesize free fatty acids. Trends Biotechnol 30: 659–667. doi: 10.1016/j.tibtech.2012.09.006
|
| [67] |
Chen L, Zhang J, Lee J, et al. (2014) Enhancement of free fatty acid production in Saccharomyces cerevisiae by control of fatty acyl-CoA metabolism. Appl Microbiol Biotechnol 98: 6739–6750. doi: 10.1007/s00253-014-5758-8
|
| [68] |
Reiser S, Somerville C (1997) Isolation of mutants of Acinetobacter calcoaceticus deficient in wax ester synthesis and complementation of one mutation with a gene encoding a fatty acyl coenzyme A reductase. J Bacteriol 179: 2969–2975. doi: 10.1128/jb.179.9.2969-2975.1997
|
| [69] |
Willis RM, Wahlen BD, Seefeldt LC, et al. (2011) Characterization of a fatty acyl-CoA reductase from Marinobacter aquaeolei VT8: a bacterial enzyme catalyzing the reduction of fatty acyl-CoA to fatty alcohol. Biochemistry 50: 10550–10558. doi: 10.1021/bi2008646
|
| [70] |
Tang X, Chen WN (2015) Enhanced production of fatty alcohols by engineering the TAGs synthesis pathway in Saccharomyces cerevisiae. Biotechnol Bioeng 112: 386–392. doi: 10.1002/bit.25356
|
| [71] |
Akhtar MK, Turner NJ, Jones PR (2013) Carboxylic acid reductase is a versatile enzyme for the conversion of fatty acids into fuels and chemical commodities. Proc Natl Acad Sci USA 110: 87–92. doi: 10.1073/pnas.1216516110
|
| [72] |
Jin Z, Wong A, Foo JL, et al. (2016) Engineering Saccharomyces cerevisiae to produce odd chain-length fatty alcohols. Biotechnol Bioeng 113: 842–851. doi: 10.1002/bit.25856
|
| [73] |
Li N, Chang WC, Warui DM, et al. (2012) Evidence for only oxygenative cleavage of aldehydes to alk(a/e)nes and formate by Cyanobacterial aldehyde decarbonylases. Biochemistry 51: 7908–7916. doi: 10.1021/bi300912n
|
| [74] | Foo JL, Susanto AV, Keasling JD, et al. (2016) Whole-cell biocatalytic and de novo production of alkanes from free fatty acids in Saccharomyces cerevisiae. Biotechnol Bioeng. |
| [75] |
Bernard A, Domergue F, Pascal S, et al. (2012) Reconstitution of plant alkane biosynthesis in Yeast demonstrates that Arabidopsis ECERIFERUM1 and ECERIFERUM3 are core components of a very-long-chain alkane synthesis complex. Plant Cell 24: 3106–3118. doi: 10.1105/tpc.112.099796
|
| [76] |
Liu JF, Nie KL, Fan LH, et al. (2013) Increased production of FAEEs for biodiesel with lipase enhanced Saccharomyces cerevisiae. Process Biochem 48: 1212–1215. doi: 10.1016/j.procbio.2013.06.003
|
| [77] |
Eriksen DT, HamediRad M, Yuan Y, et al. (2015) Orthogonal fatty acid biosynthetic pathway improves fatty acid ethyl ester production in Saccharomyces cerevisiae. ACS Synth Biol 4: 808–814. doi: 10.1021/sb500319p
|
| [78] |
Chen B, Lee DY, Chang MW (2015) Combinatorial metabolic engineering of Saccharomyces cerevisiae for terminal alkene production. Metab Eng 31: 53–61. doi: 10.1016/j.ymben.2015.06.009
|
| [79] |
Molina Grima E, Belarbi EH, Acién Fernández FG, et al. (2003) Recovery of microalgal biomass and metabolites: process options and economics. Biotechnol Adv 20: 491–515. doi: 10.1016/S0734-9750(02)00050-2
|
| [80] | Leber C, Da SN (2014) Engineering of Saccharomyces cerevisiae for the synthesis of short chain fatty acids. Biotechnol Bioeng 111: 347–358. |
| [81] |
Scharnewski M, Pongdontri P, Mora G, et al. (2008) Mutants of Saccharomyces cerevisiae deficient in acyl-CoA synthetases secrete fatty acids due to interrupted fatty acid recycling. FEBS J 275: 2765–2778. doi: 10.1111/j.1742-4658.2008.06417.x
|
| [82] |
Runguphan W, Keasling JD (2014) Metabolic engineering of Saccharomyces cerevisiae for production of fatty acid-derived biofuels and chemicals. Metab Eng 21: 103–113. doi: 10.1016/j.ymben.2013.07.003
|
| [83] |
Li X, Guo D, Cheng Y, et al. (2014) Overproduction of fatty acids in engineered Saccharomyces cerevisiae. Biotechnol Bioeng 111: 1841–1852. doi: 10.1002/bit.25239
|
| [84] |
Leber C, Polson B, Fernandez-Moya R, et al. (2015) Overproduction and secretion of free fatty acids through disrupted neutral lipid recycle in Saccharomyces cerevisiae. Metab Eng 28: 54–62. doi: 10.1016/j.ymben.2014.11.006
|
| [85] |
Kamisaka Y, Noda N, Tomita N, et al. (2006) Identification of genes affecting lipid content using transposon mutagenesis in Saccharomyces cerevisiae. Biosci Biotechnol Biochem 70: 646–653. doi: 10.1271/bbb.70.646
|
| [86] |
Kamisaka Y, Tomita N, Kimura K, et al. (2007) DGA1(diacylglycerol acyltransferase gene) overexpression and leucine biosynthesis significantly increase lipid accumulation in the Δsnf2 disruptant of Saccharomyces cerevisiae. Biochem J 408: 61–68. doi: 10.1042/BJ20070449
|
| [87] |
Kamisaka Y, Kimura K, Uemura H, et al. (2013) Overexpression of the active diacylglycerol acyltransferase variant transforms Saccharomyces cerevisiae into an oleaginous yeast. Appl Microbiol Biotechnol 97: 7345–7355. doi: 10.1007/s00253-013-4915-9
|
| [88] |
Qiao K, Imam Abidi SH, Liu H, et al. (2015) Engineering lipid overproduction in the oleaginous yeast Yarrowia lipolytica. Metab Eng 29: 56–65. doi: 10.1016/j.ymben.2015.02.005
|
| [89] |
Friedlander J, Tsakraklides V, Kamineni A, et al. (2016) Engineering of a high lipid producing Yarrowia lipolytica strain. Biotechnol Biofuels 9: 1–12. doi: 10.1186/s13068-015-0423-8
|
| [90] |
Rutter CD, Zhang S, Rao CV (2015) Engineering Yarrowia lipolytica for production of medium-chain fatty acids. Appl Microbiol Biotechnol 99: 7359–7368. doi: 10.1007/s00253-015-6764-1
|
| [91] |
Zhang S, Ito M, Skerker JM, et al. (2016) Metabolic engineering of the oleaginous yeast Rhodosporidium toruloides IFO0880 for lipid overproduction during high-density fermentation. Appl Microbiol Biotechnol 100: 9393–9405. doi: 10.1007/s00253-016-7815-y
|
| [92] |
Teo WS, Ling H, Yu AQ, et al. (2015) Metabolic engineering of Saccharomyces cerevisiae for production of fatty acid short- and branched-chain alkyl esters biodiesel. Biotechnol Biofuels 8: 1–9. doi: 10.1186/s13068-014-0179-6
|
| [93] | Yan J, Yan Y, Madzak C, et al. (2015) Harnessing biodiesel-producing microbes: from genetic engineering of lipase to metabolic engineering of fatty acid biosynthetic pathway. Crit Rev Biotechnol: 1–11. |
| [94] |
Shi S, ValleRodríguez JO, Khoomrung S, et al. (2012) Functional expression and characterization of five wax ester synthases in Saccharomyces cerevisiae and their utility for biodiesel production. Biotechnol Biofuels 5: 7–7. doi: 10.1186/PREACCEPT-1932279820621895
|
| [95] |
ValleRodríguez JO, Shi S, Siewers V, et al. (2014) Metabolic engineering of Saccharomyces cerevisiae for production of fatty acid ethyl esters, an advanced biofuel, by eliminating non-essential fatty acid utilization pathways. Appl Energy 115: 226–232. doi: 10.1016/j.apenergy.2013.10.003
|
| [96] |
Shi S, Valle-Rodríguez JO, Siewers V, et al. (2014) Engineering of chromosomal wax ester synthase integrated Saccharomyces cerevisiae mutants for improved biosynthesis of fatty acid ethyl esters. Biotechnol Bioeng 111: 1740–1747. doi: 10.1002/bit.25234
|
| [97] |
Yu KO, Jung J, Kim SW, et al. (2012) Synthesis of FAEEs from glycerol in engineered Saccharomyces cerevisiae using endogenously produced ethanol by heterologous expression of an unspecific bacterial acyltransferase. Biotechnol Bioeng 109: 110–115. doi: 10.1002/bit.23311
|
| [98] |
Thompson RA, Trinh CT (2014) Enhancing fatty acid ethyl ester production in Saccharomyces cerevisiae through metabolic engineering and medium optimization. Biotechnol Bioeng 111: 2200–2208. doi: 10.1002/bit.25292
|
| [99] |
Fillet S, Adrio JL (2016) Microbial production of fatty alcohols. World J Microbiol Biotechnol 32: 1–10. doi: 10.1007/s11274-015-1971-6
|
| [100] |
Sheng J, Stevens J, Feng X (2016) Pathway compartmentalization in peroxisome of Saccharomyces cerevisiae to produce versatile medium chain fatty alcohols. Sci Rep 6: 26884. doi: 10.1038/srep26884
|
| [101] |
Guo W, Sheng J, Zhao H, et al. (2016) Metabolic engineering of Saccharomyces cerevisiae to produce 1-hexadecanol from xylose. Microb Cell Fact 15: 1–11. doi: 10.1186/s12934-015-0402-6
|
| [102] |
Rutter CD, Rao CV (2016) Production of 1-decanol by metabolically engineered Yarrowia lipolytica. Metab Eng 38: 139–147. doi: 10.1016/j.ymben.2016.07.011
|
| [103] |
Fillet S, Gibert J, Suárez B, et al. (2015) Fatty alcohols production by oleaginous yeast. J Ind Microbiol Biotechnol 42: 1463–1472. doi: 10.1007/s10295-015-1674-x
|
| [104] |
Schirmer A, Rude MA, Li XZ, et al. (2010) Microbial biosynthesis of alkanes. Science 329: 559–562. doi: 10.1126/science.1187936
|
| [105] |
Buijs NA, Zhou YJ, Siewers V, et al. (2015) Long-chain alkane production by the yeast Saccharomyces cerevisiae. Biotechnol Bioeng 112: 1275–1279. doi: 10.1002/bit.25522
|
| [106] |
Kunjapur AM, Prather KLJ (2015) Microbial engineering for aldehyde synthesis. Appl Environ Microbiol 81: 1892–1901. doi: 10.1128/AEM.03319-14
|
| [107] |
Kunjapur AM, Cervantes B, Prather KLJ (2016) Coupling carboxylic acid reductase to inorganic pyrophosphatase enhances cell-free in vitro aldehyde biosynthesis. Biochem Eng J 109: 19–27. doi: 10.1016/j.bej.2015.12.018
|
| [108] |
Moura M, Pertusi D, Lenzini S, et al. (2016) Characterizing and predicting carboxylic acid reductase activity for diversifying bioaldehyde production. Biotechnol Bioeng 113: 944–952. doi: 10.1002/bit.25860
|
| [109] |
Kocharin K, Chen Y, Siewers V, et al. (2012) Engineering of acetyl-CoA metabolism for the improved production of polyhydroxybutyrate in Saccharomyces cerevisiae. AMB Express 2: 1–11. doi: 10.1186/2191-0855-2-1
|
| [110] |
Sanchez AM, Andrews J, Hussein I, et al. (2006) Effect of overexpression of a soluble pyridine nucleotide transhydrogenase (UdhA) on the production of poly(3-hydroxybutyrate) in Escherichia coli. Biotechnol Prog 22: 420–425. doi: 10.1021/bp050375u
|
| [111] |
Park SJ, Choi JI, Lee SY (2005) Engineering of Escherichia coli fatty acid metabolism for the production of polyhydroxyalkanoates. Enzyme Microb Technol 36: 579–588. doi: 10.1016/j.enzmictec.2004.12.005
|
| [112] |
Poirier Y, Erard N, Petétot JM (2001) Synthesis of polyhydroxyalkanoate in the peroxisome of Saccharomyces cerevisiae by using intermediates of fatty acid β-Oxidation. Appl Environ Microbiol 67: 5254–5260. doi: 10.1128/AEM.67.11.5254-5260.2001
|
| [113] |
Yazawa H, Iwahashi H, Kamisaka Y, et al. (2010) Improvement of polyunsaturated fatty acids synthesis by the coexpression of CYB5 with desaturase genes in Saccharomyces cerevisiae. Appl Microbiol Biotechnol 87: 2185–2193. doi: 10.1007/s00253-010-2679-z
|
| [114] |
Gomes A, Couto D, Alves A, et al. (2012) Trihydroxyflavones with antioxidant and anti-inflammatory efficacy. Biofactors 38: 378–386. doi: 10.1002/biof.1033
|
| [115] |
Miran SG, Lawson JE, Reed LJ (1993) Characterization of PDHβ1, the structural gene for the pyruvate dehydrogenase β subunit from Saccharomyces cerevisiae. Proc Natl Acad Sci USA 90: 1252–1256. doi: 10.1073/pnas.90.4.1252
|
| [116] |
Kratzer S, Schüller HJ (1995) Carbon source-dependent regulation of the acetyl-coenzyme A synthetase-encoding gene ACSI from Saccharomyces cerevisiae. Gene 161: 75–79. doi: 10.1016/0378-1119(95)00289-I
|
| [117] |
Ratledge C, Wynn JP (2002) The biochemistry and molecular biology of lipid accumulation in oleaginous microorganisms. Adv Appl Microbiol 51: 1–51. doi: 10.1016/S0065-2164(02)51000-5
|
| [118] |
Vemuri GN, Eiteman MA, McEwen JE, et al. (2007) Increasing NADH oxidation reduces overflow metabolism in Saccharomyces cerevisiae. Proc Natl Acad Sci USA 104: 2402–2407. doi: 10.1073/pnas.0607469104
|
| [119] |
Shiba Y, Paradise EM, Kirby J, et al. (2007) Engineering of the pyruvate dehydrogenase bypass in Saccharomyces cerevisiae for high-level production of isoprenoids. Metab Eng 9: 160–168. doi: 10.1016/j.ymben.2006.10.005
|
| [120] |
Chen Y, Daviet L, Schalk M, et al. (2013) Establishing a platform cell factory through engineering of yeast acetyl-CoA metabolism. Metab Eng 15: 48–54. doi: 10.1016/j.ymben.2012.11.002
|
| [121] |
Krivoruchko A, Serrano-Amatriain C, Chen Y, et al. (2013) Improving biobutanol production in engineered Saccharomyces cerevisiae by manipulation of acetyl-CoA metabolism. J Ind Microbiol Biotechnol 40: 1051–1056. doi: 10.1007/s10295-013-1296-0
|
| [122] |
Tang X, Feng H, Chen WN (2013) Metabolic engineering for enhanced fatty acids synthesis in Saccharomyces cerevisiae. Metab Eng 16: 95–102. doi: 10.1016/j.ymben.2013.01.003
|
| [123] | Shi S, Chen Y, Siewers V, et al. (2014) Improving production of malonyl coenzyme a-derived metabolites by abolishing Snf1-dependent regulation of Acc1. mBio 5: e01130–01114. |
| [124] |
Wang Y, Chen H, Yu O (2014) A plant malonyl-CoA synthetase enhances lipid content and polyketide yield in yeast cells. Appl Microbiol Biotechnol 98: 5435–5447. doi: 10.1007/s00253-014-5612-z
|
| [125] | Zhang Y, Adams IP, Ratledge C (2007) Malic enzyme: the controlling activity for lipid production? Overexpression of malic enzyme in Mucor circinelloides leads to a 2.5-fold increase in lipid accumulation. Microbiology 153: 2013–2025. |
| [126] |
Wynn JP, Ratledge C (1997) Malic enzyme is a major source of NADPH for lipid accumulation by Aspergillus Nidulans. Microbiology 143: 253–257. doi: 10.1099/00221287-143-1-253
|
| [127] |
Minard KI, McAlisterhenn L (2005) Sources of NADPH in Yeast vary with carbon source. J Biol Chem 280: 39890–39896. doi: 10.1074/jbc.M509461200
|
| [128] |
Grabowska D, Chelstowska A (2003) The ALD6 gene product is indispensable for providing NADPH in Yeast cells lacking glucose-6-phosphate dehydrogenase activity. J Biol Chem 278: 13984–13988. doi: 10.1074/jbc.M210076200
|
| [129] | Sheppard MJ, Kunjapur AM, Wenck SJ, et al. (2014) Retro-biosynthetic screening of a modular pathway design achieves selective route for microbial synthesis of 4-methyl-pentanol. Nat Commun 5: 1–10. |
| [130] |
Andre C, Kim SW, Yu XH, et al. (2013) Fusing catalase to an alkane-producing enzyme maintains enzymatic activity by converting the inhibitory byproduct H2O2 to the cosubstrate O2. Proc Natl Acad Sci USA 110: 3191–3196. doi: 10.1073/pnas.1218769110
|
| [131] |
Khara B, Menon N, Levy C, et al. (2013) Production of propane and other short-chain alkanes by structure-based engineering of ligand specificity in aldehyde-deformylating oxygenase. Chembiochem 14: 1204–1208. doi: 10.1002/cbic.201300307
|
| [132] |
Wang Q, Huang X, Zhang J, et al. (2014) Engineering self-sufficient aldehyde deformylating oxygenases fused to alternative electron transfer systems for efficient conversion of aldehydes into alkanes. Chem Commun 50: 4299–4301. doi: 10.1039/c4cc00591k
|
| [133] |
Lo TM, Teo WS, Ling H, et al. (2013) Microbial engineering strategies to improve cell viability for biochemical production. Biotechnol Adv 31: 903–914. doi: 10.1016/j.biotechadv.2013.02.001
|
| [134] | de Jong BW, Siewers V, Nielsen J (2015) Physiological and transcriptional characterization of Saccharomyces cerevisiae engineered for production of fatty acid ethyl esters. FEMS Yeast Res 16. |
| [135] |
Chen L, Lee JJL, Zhang J, et al. (2016) Comparative proteomic analysis of engineered Saccharomyces cerevisiae with enhanced free fatty acid accumulation. Appl Microbiol Biotechnol 100: 1407–1420. doi: 10.1007/s00253-015-7028-9
|
| [136] |
Ling H, Chen B, Kang A, et al. (2013) Transcriptome response to alkane biofuels in Saccharomyces cerevisiae: identification of efflux pumps involved in alkane tolerance. Biotechnol Biofuels 6: 1–10. doi: 10.1186/1754-6834-6-1
|
| [137] |
Chen B, Ling H, Chang MW (2013) Transporter engineering for improved tolerance against alkane biofuels in Saccharomyces cerevisiae. Biotechnol Biofuels 6: 1–10. doi: 10.1186/1754-6834-6-1
|
| [138] |
Ling H, Pratomo Juwono NK, Teo WS, et al. (2015) Engineering transcription factors to improve tolerance against alkane biofuels in Saccharomyces cerevisiae. Biotechnol Biofuels 8: 1–11. doi: 10.1186/s13068-014-0179-6
|
| [139] |
Rodriguez GM, Atsumi S (2014) Toward aldehyde and alkane production by removing aldehyde reductase activity in Escherichia coli. Metab Eng 25: 227–237. doi: 10.1016/j.ymben.2014.07.012
|
| [140] |
Xu P, Li L, Zhang F, et al. (2014) Improving fatty acids production by engineering dynamic pathway regulation and metabolic control. Proc Natl Acad Sci USA 111: 11299–11304. doi: 10.1073/pnas.1406401111
|
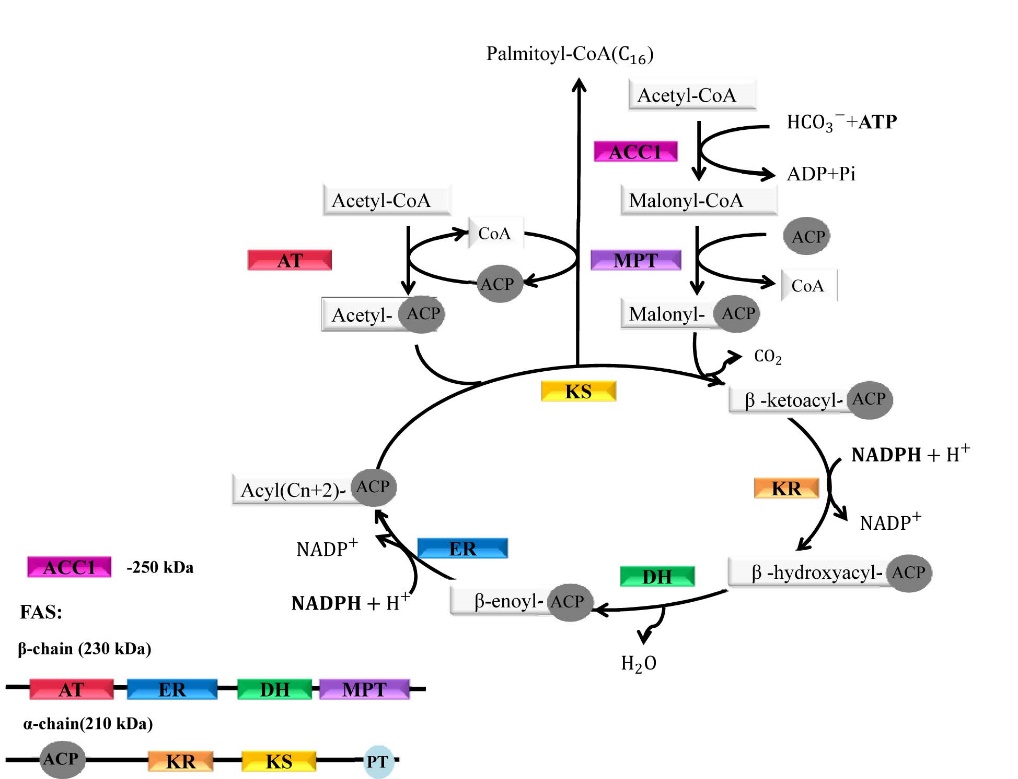
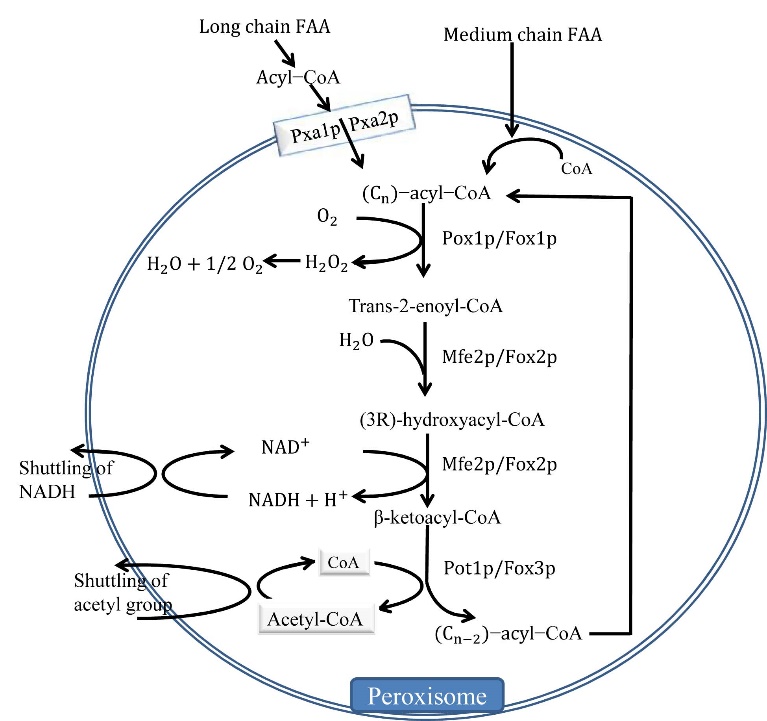
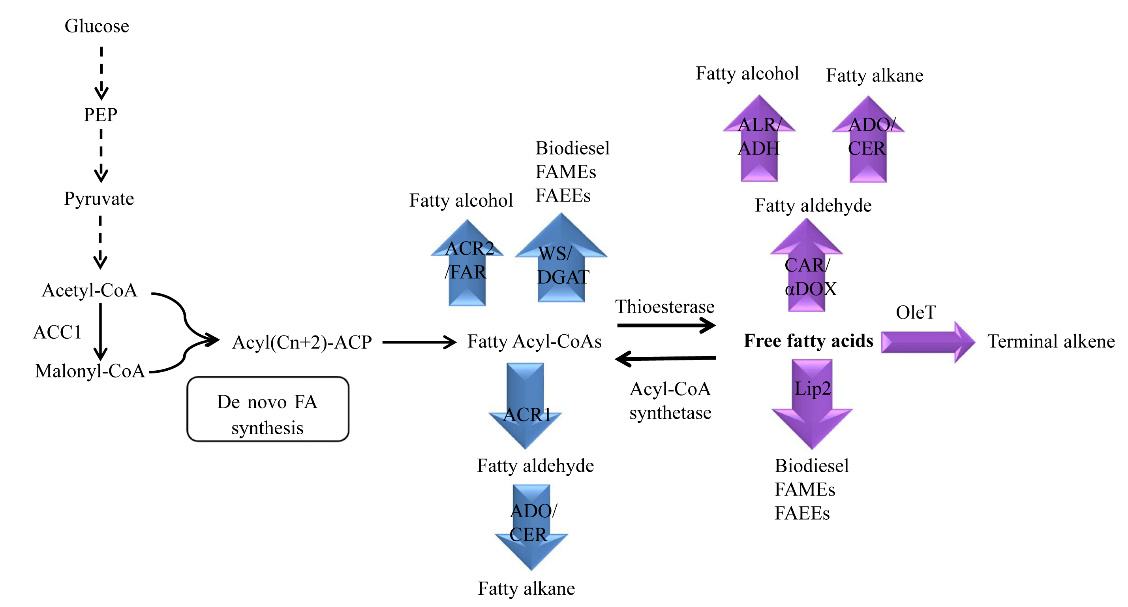
Figures(3)

Liwei Chen, Jaslyn Lee, Wei Ning Chen. The use of metabolic engineering to produce fatty acid-derived biofuel and chemicals in Saccharomyces cerevisiae: a review[J]. AIMS Bioengineering, 2016, 3(4): 468-492. doi: 10.3934/bioeng.2016.4.468







 DownLoad:
DownLoad: