Citation: Shan-Yang Lin. Salmon calcitonin: conformational changes and stabilizer effects[J]. AIMS Biophysics, 2015, 2(4): 695-723. doi: 10.3934/biophy.2015.4.695

| [1] |
Fosgerau K, Hoffmann T (2015) Peptide therapeutics: current status and future directions. Drug Discov Today 20: 122-128. doi: 10.1016/j.drudis.2014.10.003

|
| [2] |
Leader B, Baca QJ, Golan DE (2008) Protein therapeutics: a summary and pharmacological classification. Nat Rev Drug Discov 7: 21-39. doi: 10.1038/nrd2399
|
| [3] |
Craik DJ, Fairlie DP, Liras S, et al. (2013) The future of peptide-based drugs. Chem Biol Drug Des 81: 136-147. doi: 10.1111/cbdd.12055
|
| [4] |
Bruno BJ, Miller GD, Lim CS (2013) Basics and recent advances in peptide and protein drug delivery. Ther Deliv 4: 1443-1467. doi: 10.4155/tde.13.104
|
| [5] | Ratnaparkhi MP, Chaudhari SP, Pandya VA (2011) Peptides and proteins in pharmaceuticals. Int J Curr Pharm Res 3 (2): 1-9. |
| [6] |
Mullard A (2011) 2010 FDA Approvals. Nat Rev Drug Discov 10: 82-85. doi: 10.1038/nrd3370
|
| [7] |
Kneller R (2010) The importance of new companies for drug discovery: Origins of a decade of new drugs. Nat Rev Drug Discov 9: 867-882. doi: 10.1038/nrd3251
|
| [8] |
Uhlig T, Kyprianou TD, Martinelli FG, et al. (2014) The emergence of peptides in the pharmaceutical business: From exploration to exploitation. EuPA Open Proteom 4: 58-69. doi: 10.1016/j.euprot.2014.05.003
|
| [9] |
Frokjaer S, Otzen DE (2005) Protein drug stability: a formulation challenge. Nat Rev Drug Discov 4: 298-306. doi: 10.1038/nrd1695
|
| [10] | van de Weert M, Randolph TW (2012) Physical instability of peptides and pProteins, In: Hovgaard L, Frokjaer S, Van De Weert M, Pharmaceutical Formulation Development of Peptides and Proteins, 2 Eds. , Florida: CRC Press, 107-129. |
| [11] |
Rathore N, Rajan RS (2008) Current perspectives on stability of protein drug products during formulation, fill and finish operations. Biotechnol Prog 24: 504-514. doi: 10.1021/bp070462h
|
| [12] |
Taverna DM, Goldstein RA (2002) Why are proteins marginally stable? Proteins 46: 105-109. doi: 10.1002/prot.10016
|
| [13] | Williams PD, Pollock DD, Goldstein RA (2006) Functionality and the evolution of marginal stability in proteins: Inferences from lattice simulations. Evol Bioinform Online 2: 91-101. |
| [14] | Chang BS, Yeung B (2010) Physical stability of protein pharmaceuticals, In: Jameel F, Hershenson S, Formulation and Process Development Strategies for Manufacturing Biopharmaceuticals, New Jersey: John Wiley & Sons Inc, 69-104. |
| [15] |
Lai MC, Topp EM (1999) Solid-state chemical stability of proteins and peptides. J Pharm Sci 88: 489-500. doi: 10.1021/js980374e
|
| [16] |
Chaudhuri R, Cheng Y, Middaugh CR, et al. (2014) High-throughput biophysical analysis of protein therapeutics to examine interrelationships between aggregate formation and conformational stability. AAPS J 16: 48-64. doi: 10.1208/s12248-013-9539-6
|
| [17] | Jacob S, Shirwaikar A, Srinivasan K, et al. (2006) Stability of proteins in aqueous solution and solid state. Indian J Pharm Sci 68: 154-163. |
| [18] |
Manning MC, Chou DK, Murphy BM, et al. (2010) Stability of protein pharmaceuticals: an update. Pharm Res 27: 544-575. doi: 10.1007/s11095-009-0045-6
|
| [19] | Carpenter JF, Manning MC (2002) Rational Design of Stable Protein Formulations: Theory and Practice, New York: Kluwert Academic /Plenum Publishers. |
| [20] | Pace CN, Grimsley GR, Scholtz JM, et al. (2014) Protein stability. In: eLS. John Wiley & Sons Ltd, Chichester. Available from: http://onlinelibrary. wiley. com/doi/10. 1002/9780470015902. a0003002. pub3/otherversions |
| [21] | Banga AK (2015) Therapeutic Peptides and Proteins: Formulation, Processing, and Delivery Systems, 3 Eds. , Florida: CRC Press. |
| [22] | Ohtake S, Wang W (2013) Protein and peptide formulation development. Pharmaceutical Sciences Encyclopedia 11: 1-44. |
| [23] |
Krishnamurthy R, Manning MC (2002) The stability factor: importance in formulation development. Curr Pharm Biotechnol 3: 361-371. doi: 10.2174/1389201023378229
|
| [24] | Cicerone MT, Pikal MJ, Qian KK (2015) Stabilization of proteins in solid form. Adv Drug Deliv Rev doi: 10. 1016/j. addr. 2015. 05. 006. |
| [25] | Murphy KP (2001) Protein structure, stability, and folding, Series: Methods in Molecular Biology, Vol. 168, New Jersey: Humana Press. |
| [26] | Scheeff ED, Fink JL (2003) Fundamentals of protein structure. Methods Biochem Anal 44: 15-39. |
| [27] | Adler MJ, Jamieson AG, Hamilton AD (2011) Hydrogen-bonded synthetic mimics of protein secondary structure as disruptors of protein-protein interactions. Curr Top Microbiol Immunol 348: 1-23. |
| [28] | Kwok SC, Mant CT, Hodges RS (2002) Importance of secondary structural specificity determinants in protein folding: insertion of a native beta-sheet sequence into an alpha-helical coiled-coil. Protein Sci 11: 1519-1531. |
| [29] |
Jorgensen L, Hostrup S, Moeller EH, et al. (2009) Recent trends in stabilizing peptides and proteins in pharmaceutical formulation - considerations in the choice of excipients. Expert Opin Drug Deliv 6: 1219-1230. doi: 10.1517/17425240903199143
|
| [30] |
Hirsch PF, Baruch H (2003) Is calcitonin an important physiological substance? Endocrine 21: 201-208. doi: 10.1385/ENDO:21:3:201
|
| [31] | Huang CL, Sun L, Moonga BS, et al. (2006) Molecular physiology and pharmacology of calcitonin. Cell Mol Biol 52: 33-43. |
| [32] |
Väänänen K (2005) Mechanism of osteoclast mediated bone resorption--rationale for the design of new therapeutics. Adv Drug Deliv Rev 57: 959-971. doi: 10.1016/j.addr.2004.12.018
|
| [33] |
Davey RA, Findlay DM (2013) Calcitonin: physiology or fantasy? J Bone Miner Res 28: 973-979. doi: 10.1002/jbmr.1869
|
| [34] |
Felsenfeld AJ, Levine BS (2015) Calcitonin, the forgotten hormone: does it deserve to be forgotten? Clin Kidney J 8: 180-187. doi: 10.1093/ckj/sfv011
|
| [35] | Pondel M (2000) Calcitonin and calcitonin receptors: bone and beyond. Int J Exp Pathol 81: 405-422. |
| [36] | Endres DB, Rude RK (1999) Mineral and bone metabolism, In: Burtis CA, Ashwood ER Author, Textbook of Clinical Chemistry, 3 Eds. , Pennsylvania: W B Saunders Company, 1395-1457 |
| [37] | Ramasamy I (2006) Recent advances in physiological calcium homeostasis. Clin Chem Lab Med 44: 237-273. |
| [38] |
Chesnut CH 3rd, Azria M, Silverman S, et al. (2008) Salmon calcitonin: a review of current and future therapeutic indications. Osteoporos Int 19: 479-491. doi: 10.1007/s00198-007-0490-1
|
| [39] |
Chakraborty C, Nandi S, Sinha S (2004) Overexpression, purification and characterization of recombinant salmon calcitonin, a therapeutic protein, in Streptomyces avermitilis. Protein Pept Lett 11: 165-173. doi: 10.2174/0929866043478266
|
| [40] | O'Connell MB (2006) Prescription drug therapies for prevention and treatment of postmenopausal osteoporosis. J Manag Care Pharm 12(6 Suppl A): S10-9, quiz S26-8. |
| [41] | Sexton PM, Findlay DM, Martin TJ (1999) Calcitonin. Curr Med Chem 6: 1067-1093. |
| [42] |
Windich V, De Luccia F, Herman F, et al. (1997) Degradation pathways of salmon calcitonin in aqueous solution. J Pharm Sci 86: 359-364. doi: 10.1021/js9602305
|
| [43] |
Torres-Lugo M, Peppas NA (2000) Transmucosal delivery systems for calcitonin: a review. Biomaterials 21: 1191-1196. doi: 10.1016/S0142-9612(00)00011-9
|
| [44] | Azria M (2003) Osteoporosis management in day-to-day practice. The role of calcitonin. J Musculoskelet Neuronal Interact 3: 210-213. |
| [45] | Karsdal MA, Henriksen K, Arnold M , et al. (2008) Calcitonin: a drug of the past or for the future? Physiologic inhibition of bone resorption while sustaining osteoclast numbers improves bone quality. BioDrugs 22: 137-144. |
| [46] |
Azria M, Copp DH, Zanelli JM (1995) 25 years of salmon-calcitonin—from synthesis to therapeutic use. Calcif Tissue Int 57: 405-408. doi: 10.1007/BF00301940
|
| [47] |
D'Hondt M, Van Dorpe S, Mehuys E, et al. (2010) Quality analysis of salmon calcitonin in a polymeric bioadhesive pharmaceutical formulation: sample preparation optimization by DOE. J Pharm Biomed Anal 53: 939-945. doi: 10.1016/j.jpba.2010.06.028
|
| [48] |
Hong B, Wu B, Li Y (2003) Production of C-terminal amidated recombinant salmon calcitonin in Streptomyces lividans. Appl Biochem Biotechnol 110: 113-123. doi: 10.1385/ABAB:110:2:113
|
| [49] |
Andreassen KV, Hjuler ST, Furness SG, et al. (2014) Prolonged calcitonin receptor signaling by salmon, but not human calcitonin, reveals ligand bias. PLoS One 9: e92042. doi: 10.1371/journal.pone.0092042
|
| [50] |
Stevenson JC, Evans IM (1981) Pharmacology and therapeutic use of calcitonin. Drugs 21: 257-272. doi: 10.2165/00003495-198121040-00002
|
| [51] |
Renukuntla J, Vadlapudi AD, Patel A, et al. (2013) Approaches for enhancing oral bioavailability of peptides and proteins. Int J Pharm 447: 75-93. doi: 10.1016/j.ijpharm.2013.02.030
|
| [52] |
Hoyer H, Perera G, Bernkop-Schnürch A (2010) Noninvasive delivery systems for peptides and proteins in osteoporosis therapy: a retroperspective. Drug Dev Ind Pharm 36: 31-44. doi: 10.3109/03639040903059342
|
| [53] | Satoh T, Yoshida G, Orito Y, et al. (1998) Drug delivery system for the treatment of osteoporosis. Nihon Rinsho 56: 742-747. |
| [54] | Sinsuebpol C, Chatchawalsaisin J, Kulvanich P (2013) Preparation and in vivo absorption evaluation of spray dried powders containing salmon calcitonin loaded chitosan nanoparticles for pulmonary delivery. Drug Des Devel Ther 7: 861-873. |
| [55] |
Tas C, Mansoor S, Kalluri H, et al. (2012) Delivery of salmon calcitonin using a microneedle patch. Int J Pharm 423: 257-263. doi: 10.1016/j.ijpharm.2011.11.046
|
| [56] |
Cholewinsky M, Luckel B, Horn H (1996) Degradation pathways, analytical characterization and formulation strategies of a peptide and a protein Calcitonin and human growth hormone in comparison. Pharm Acta Helv 71: 405-419. doi: 10.1016/S0031-6865(96)00049-0
|
| [57] |
Uda K, Kobayashi Y, Hisada T, et al. (1999) Stable human calcitonin analogues with high potency on bone together with reduced anorectic and renal actions. Biol Pharm Bull 22: 244-252. doi: 10.1248/bpb.22.244
|
| [58] |
Cudd A, Arvinte T, Das RE, et al. (1995) Enhanced potency of human calcitonin when fibrillation is avoided. J Pharm Sci 84: 717-719. doi: 10.1002/jps.2600840610
|
| [59] | Wang W, Roberts CJ (2010) Aggregation of Therapeutic Proteins, 1 Eds. , New Jersey: John Wiley & Sons Inc. |
| [60] |
den Engelsman J, Garidel P, Smulders R, et al. (2011) Strategies for the assessment of protein aggregates in pharmaceutical biotech product development. Pharm Res 28: 920-933. doi: 10.1007/s11095-010-0297-1
|
| [61] | Bryan J (2014) Protein aggregation: formulating a problem. Pharm J 293: 7826. |
| [62] |
Mahler HC, Friess W, Grauschopf U, et al. (2009) Protein aggregation: pathways, induction factors, and analysis. J Pharm Sci 98: 2909-2934. doi: 10.1002/jps.21566
|
| [63] |
Brange J, Andersen L, Laursen ED, et al. (1997) Toward understanding insulin fibrillation. J Pharm Sci 86: 517-525. doi: 10.1021/js960297s
|
| [64] |
Shire SJ, Shahrokh Z, Liu J (2004) Challenges in the development of high protein concentration formulations. J Pharm Sci 93: 1390-1402. doi: 10.1002/jps.20079
|
| [65] |
Thirumangalathu R, Krishnan S, Brems DN, et al. (2006) Effects of pH, temperature, and sucrose on benzyl alcohol-induced aggregation of recombinant human granulocyte colony stimulating factor. J Pharm Sci 95: 1480-1497. doi: 10.1002/jps.20619
|
| [66] |
Wang W, Wang YJ, Wang DQ (2008) Dual effects of Tween 80 on protein stability. Int J Pharm 347: 31-38. doi: 10.1016/j.ijpharm.2007.06.042
|
| [67] |
Carpenter JF, Pikal MJ, Chang BS, et al. (1997) Rational design of stable lyophilized protein formulations: Some practical advice. Pharm Res 14: 969-975. doi: 10.1023/A:1012180707283
|
| [68] |
Chi EY, Weickmann J, Carpenter JF, et al. (2005) Heterogeneous nucleation-controlled particulate formation of recombinant human platelet-activating factor acetylhydrolase in pharmaceutical formulation. J Pharm Sci 94: 256-274. doi: 10.1002/jps.20237
|
| [69] |
KatakamM, Bell LN, Banga AK (1995) Effect of surfactants on the physical stability of recombinant human growth hormone. J Pharm Sci 84: 713-716. doi: 10.1002/jps.2600840609
|
| [70] |
Taylor JW, Jin QK, Sbacchi M, et al. (2002) Side-chain lactam-bridge conformational constraints differentiate the activities of salmon and human calcitonins and reveal a new design concept for potent calcitonin analogues. J Med Chem 45: 1108-1121. doi: 10.1021/jm010474o
|
| [71] | Karsdal MA, Henriksen K, Arnold M, et al. (2008) Calcitonin: a drug of the past or for the future? Physiologic inhibition of bone resorption while sustaining osteoclast numbers improves bone quality. BioDrugs 22: 137-144. |
| [72] | Kapurniotu A, Taylor JW (1995) Structural and conformational requirements for human calcitonin activity: Design, synthesis, and study of lactam-bridged Analogues. J Med Chem 3: 836-847. |
| [73] | Cholewinski M, Lückel B, Horn H. (1996) Degradation pathways, analytical characterization and formulation strategies of a peptide and a protein calcitonine and human growth hormone in comparison. Pharm Acta Helv 71: 405-419. |
| [74] | Wang W (1999) Instability, stabilization, and formulation of liquid protein pharmaceuticals. Int J Pharm 185: 129-188. |
| [75] | Wang W (2000) Lyophilization and development of solid protein pharmaceuticals. Int J Pharm 203: 1-60. |
| [76] |
Rathore N, Rajan RS (2008) Current perspectives on stability of protein drug products during formulation, fill and finish operations. Biotechnol Prog 24: 504-514. doi: 10.1021/bp070462h
|
| [77] |
Krishnamurthy R, Manning MC (2002) The stability factor: importance in formulation development. Curr Pharm Biotechnol 3: 361-371. doi: 10.2174/1389201023378229
|
| [78] |
Chang SL, Hofmann GA, Zhang L, et al. (2003) Stability of a transdermal salmon calcitonin formulation. Drug Deliv 10: 41-45. doi: 10.1080/713840326
|
| [79] |
Bauer HH, Aebi U, Häner M, et al. (1995) Architecture and polymorphism of fibrillar supramolecular assemblies produced by in vitro aggregation of human calcitonin. J Struct Biol 115: 1-15. doi: 10.1006/jsbi.1995.1024
|
| [80] |
Avidan-Shpalter C, Gazit E (2006) The early stages of amyloid formation: biophysical and structural characterization of human calcitonin pre-fibrillar assemblies. Amyloid 13: 216-225. doi: 10.1080/13506120600960643
|
| [81] |
Diociaiuti M, Gaudiano MC, Malchiodi-Albedi F (2011) The slowly aggregating salmon Calcitonin: a useful tool for the study of the amyloid oligomers structure and activity. Int J Mol Sci 12: 9277-9295. doi: 10.3390/ijms12129277
|
| [82] |
Seyferth S, Lee G (2003) Structural studies of EDTA-induced fibrillation of salmon calcitonin. Pharm Res 20: 73-80. doi: 10.1023/A:1022250809235
|
| [83] |
Nakamuta H, Orlowski RC, Epand RM (1990) Evidence for calcitonin receptor heterogenecity: binding studies with non-helical analogs. Endocrinology 127: 163-169. doi: 10.1210/endo-127-1-163
|
| [84] |
Siligardi G, Samori B, Melandri S, et al. (1994) Correlations between biological activities and conformation properties for human, salmon, eel, porcine calcitonins and Elcatonin elucidated by CD spectroscopy. Eur J Biochem 221: 1117-1125. doi: 10.1111/j.1432-1033.1994.tb18832.x
|
| [85] |
Moriarty DF, Vagts S, Raleigh DP (1998) A role for the C-terminus of calcitonin in aggregation and gel formation: a comparative study of C-terminal Fragments of human and salmon calcitonin. Biochem Biophys Res Commun 245: 344-348. doi: 10.1006/bbrc.1998.8425
|
| [86] |
van Dijkhuizen-Radersma R, Nicolas HM, van de Weert M, et al. (2002) Stability aspects of salmon calcitonin entrapped in poly(ether-ester) sustained release systems. Int J Pharm 248: 229-237. doi: 10.1016/S0378-5173(02)00458-1
|
| [87] |
Tang Y, Singh J (2010) Thermosensitive drug delivery system of salmon calcitonin: in vitro release, in vivo absorption, bioactivity and therapeutic efficacies. Pharm Res 27: 272-284. doi: 10.1007/s11095-009-0015-z
|
| [88] |
Windisch V, Deluccia F, Duhau L, et al. (1997) Degradation pathways of salmon calcitonin in aqueous solution. J Pharm Sci 86: 359-364. doi: 10.1021/js9602305
|
| [89] |
Lucke A, Kiermaier J, Gopferich A (2002) Peptide acylation by poly(α-hydroxy esters). Pharm Res 19: 175-181. doi: 10.1023/A:1014272816454
|
| [90] |
Montgomerie S, Sundararaj S, Gallin WJ, et al. (2006) Improving the accuracy of protein secondary structure prediction using structural alignment. BMC Bioinformatics 7: 301. doi: 10.1186/1471-2105-7-301
|
| [91] | Rost B (2001) Review: protein secondary structure prediction continues to rise. J Struct Biol 134: 204-218. |
| [92] |
Pirovano W, Heringa J (2010) Protein secondary structure prediction. Methods Mol Biol 609: 327-348. doi: 10.1007/978-1-60327-241-4_19
|
| [93] |
Kwok SC, Mant CT, Hodges RS (2002) Importance of secondary structural specificity determinants in protein folding: insertion of a native beta-sheet sequence into an alpha-helical coiled-coil. Protein Sci 11: 1519-1531. doi: 10.1110/ps.4170102
|
| [94] |
Ji YY, Li YQ (2010) The role of secondary structure in protein structure selection. Eur Phys J E Soft Matter 32: 103-107. doi: 10.1140/epje/i2010-10591-5
|
| [95] |
Haris PI, Chapman D (1995) The conformational analysis of peptides using Fourier transform IR spectroscopy. Biopolymers 37: 251-263. doi: 10.1002/bip.360370404
|
| [96] |
Manning MC (2005) Use of infrared spectroscopy to monitor protein structure and stability. Expert Rev Proteomics 2: 731-743. doi: 10.1586/14789450.2.5.731
|
| [97] | Kong J, Yu S (2007) Fourier transform infrared spectroscopic analysis of protein secondary structures. Acta Biochim Biophys Sinica 39: 549-559 |
| [98] |
Carpenter JF, Chang BS, Garzon-Rodriguez W, et al. (2002) Rational design of stable lyophilized protein formulations: Theory and practice. Pharm Biotechnol 13: 109-133. doi: 10.1007/978-1-4615-0557-0_5
|
| [99] |
Chang LL, Pikal MJ (2009) Mechanisms of protein stabilization in the solid state. J Pharm Sci 98: 2886-2908. doi: 10.1002/jps.21825
|
| [100] |
Wang W (2000) Lyophilization and development of solid protein pharmaceuticals. Int J Pharm 203: 1-60. doi: 10.1016/S0378-5173(00)00423-3
|
| [101] |
Maa Y-F, Prestrelski SJ (2000) Biopharmaceutical powders: Particle formation and formulation considerations. Curr Pharm Biotechnol 1: 283-302. doi: 10.2174/1389201003378898
|
| [102] |
Abdul-Fattah AM, Kalonia DS, Pikal MJ (2007) The challenge of drying method selection for protein pharmaceuticals: Product quality implications. J Pharm Sci 96: 1886-1916. doi: 10.1002/jps.20842
|
| [103] |
Abdul-Fattah AM, Truong-Le V, Yee L, et al. (2007) Drying induced variations in physico-chemical properties of amorphous pharmaceuticals and their impact on stability (I): Stability of a monoclonal antibody. J Pharm Sci 96: 1983-2008. doi: 10.1002/jps.20859
|
| [104] |
Klibanov AM, Schefiliti JA (2004) On the relationship between conformation and stability in solid pharmaceutical protein formulations. Biotechnol Lett 26: 1103-1106. doi: 10.1023/B:BILE.0000035520.47933.a6
|
| [105] |
Angkawinitwong U, Sharma G, Khaw PT, et al. (2015) Solid-state protein formulations. Ther Deliv 6: 59-82. doi: 10.4155/tde.14.98
|
| [106] |
Lee TH, Cheng WT, Lin SY (2010) Thermal stability and conformational structure of salmon calcitonin in the solid and liquid states. Biopolymers 93: 200-207. doi: 10.1002/bip.21323
|
| [107] |
Stevenson CL, Tan MM (2000) Solution stability of salmon calcitonin at high concentration for delivery in an implantable system. J Pept Res 55: 129-139. doi: 10.1034/j.1399-3011.2000.00160.x
|
| [108] |
Dong A, Huang P, Caughey WS (1990) Protein secondary structures in water from second-derivative amide I infrared spectra. Biochemistry 29: 3303-3308. doi: 10.1021/bi00465a022
|
| [109] | Kanari K, Nosaka A (1995) Study of human calcitonin fibrillation by proton nuclear magnetic resonance spectroscopy. Biochemistry 34: 12138-12143. |
| [110] | Arvinte T, Drake A (1993) Comparative study of human and salmon calcitonin secondary structure in solutions with low dielectric constants. J Biol Chem 268: 6408-6414. |
| [111] |
Lee SL, Yu LX, Cai B, et al. (2011) Scientific considerations for generic synthetic salmon calcitonin nasal spray products. AAPS J 13: 14-19. doi: 10.1208/s12248-010-9242-9
|
| [112] |
Binkley N, Bone H, Gilligan JP, et al. (2014) Efficacy and safety of oral recombinant calcitonin tablets in postmenopausal women with low bone mass and increased fracture risk: a randomized, placebo-controlled trial. Osteoporos Int 25: 2649-2656. doi: 10.1007/s00198-014-2796-0
|
| [113] |
Murphy LR, Matubayasi N, Payne VA, et al. (1998) Protein hydration and unfolding-insights from experimental partial specific volumes and unfolded protein models. Fold Des 3: 105-118. doi: 10.1016/S1359-0278(98)00016-9
|
| [114] |
Schiffer CA, Dötsch V (1996) The role of protein-solvent interactions in protein unfolding. Curr Opin Biotechnol 7: 428-432. doi: 10.1016/S0958-1669(96)80119-4
|
| [115] |
Lee JC (2000) Biopharmaceutical formulation. Curr Opin Biotechnol 11: 81-84. doi: 10.1016/S0958-1669(99)00058-0
|
| [116] |
Canchi DR, García AE (2013) Cosolvent effects on protein stability. Annu Rev Phys Chem 64: 273-293. doi: 10.1146/annurev-physchem-040412-110156
|
| [117] |
England JL, Haran G (2011) Role of solvation effects in protein denaturation: from thermodynamics to single molecules and back. Annu Rev Phys Chem 62: 257-277. doi: 10.1146/annurev-physchem-032210-103531
|
| [118] | Lee TH, Lin SY (2011) Pluronic F68 enhanced the conformational stability of salmon calcitonin in both aqueous solution and lyophilized solid form. Biopolymers 95: 785-791. |
| [119] |
Lee TH, Lin SY (2011) Additives affecting thermal stability of salmon calcitonin in aqueous solution and structural similarity in lyophilized solid form. Process Biochem 46: 2163-2169. doi: 10.1016/j.procbio.2011.08.017
|
| [120] |
Andreotti G, Méndez BL, Amodeo P, et al. (2006) Structural determinants of salmon calcitonin bioactivity: the role of the Leu-based amphipathic alpha-helix. J Biol Chem 281: 24193-24203. doi: 10.1074/jbc.M603528200
|
| [121] |
Carpenter JF, Prestrelski SJ, Dong A (1998) Application of infrared spectroscopy to development of stable lyophilized protein formulations. Eur J Pharm Biopharm 45: 231-238. doi: 10.1016/S0939-6411(98)00005-8
|
| [122] | Haris PI, Chapman D (1994) Analysis of polypeptide and protein structures using Fourier transform infrared spectroscopy. Methods Mol Biol 22: 183-202. |
| [123] |
Pedone E, Bartolucci S, Rossi M, et al. (2003) Structural and thermal stability analysis of Escherichia coli and Alicyclobacillus acidocaldarius thioredoxin revealed a molten globule-like state in thermal denaturation pathway of the proteins: an infrared spectroscopic study. Biochem J 373: 875-883. doi: 10.1042/bj20021747
|
| [124] | Cook TJ, Shenoy SS (2002) Stability of calcitonin salmon in nasal spray at elevated temperatures. Am J Health Syst Pharm 59: 713-715. |
| [125] |
Lee KC, Lee YJ, Song HM, et al. (1992) Degradation of synthetic salmon calcitonin in aqueous solution. Pharm Res 9: 1521-1523. doi: 10.1023/A:1015839719618
|
| [126] |
Windisch V, DeLuccia F, Duhau L, et al. (1997) Degradation pathways of salmon calcitonin in aqueous solution. J Pharm Sci 86: 359-364. doi: 10.1021/js9602305
|
| [127] |
Kamihira M, Naito A, Tuzi S, et al. (2000) Conformational transitions and fibrillation mechanism of human calcitonin as studied by high-resolution solid-state 13C NMR. Protein Sci 9: 867-877. doi: 10.1110/ps.9.5.867
|
| [128] | Andreotti G, Motta A (2004) Modulating calcitonin fibrillogenesis: an antiparallel alpha-helical dimer inhibits fibrillation of salmon calcitonin. J Biol Chem 279: 6364-6370. |
| [129] |
Gaudiano MC, Colone M, Bombelli C, et al. (2005) Early stages of salmon calcitonin aggregation: effect induced by ageing and oxidation processes in water and in the presence of model membranes. Biochim Biophys Acta 1750: 134-145. doi: 10.1016/j.bbapap.2005.04.008
|
| [130] |
Diociaiuti M, Macchia G, Paradisi S, et al. (2014) Native metastable prefibrillar oligomers are the most neurotoxic species among amyloid aggregates. Biochim Biophys Acta 1842: 1622-1629. doi: 10.1016/j.bbadis.2014.06.006
|
| [131] |
Gilchrist PJ, Bradshow JP (1993) Amyloid formation by salmon calcitonin. Biochim Biophys Acta 1182: 111-114. doi: 10.1016/0925-4439(93)90160-3
|
| [132] | Arvinte T, Cudd A, Drake AF (1993) The structure and mechanism of formation of human calcitonin fibrils. J Biol Chem 268: 6415-6422. |
| [133] |
Rastogi N, Mitra K, Kumar D, et al. (2012) Metal ions as cofactors for aggregation of therapeutic peptide salmon calcitonin. Inorg Chem 51: 5642-5650. doi: 10.1021/ic202604v
|
| [134] |
Diociaiuti M, Macchia G, Paradisi S, et al. (2014) Native metastable prefibrillar oligomers are the most neurotoxic species among amyloid aggregates. Biochim Biophys Acta 1842: 1622-1629. doi: 10.1016/j.bbadis.2014.06.006
|
| [135] |
Rawat A, Kumar D (2013) NMR investigations of structural and dynamics features of natively unstructured drug peptide-salmon calcitonin: implication to rational design of potent sCT analogs. J Pept Sci 19: 33-45. doi: 10.1002/psc.2471
|
| [136] |
Allison SD, Randolph TW, Manning MC, et al. (1998) Effects of drying methods and additives on structure and function of actin: mechanisms of dehydration-induced damage and its inhibition. Arch Biochem Biophys 358: 171-181. doi: 10.1006/abbi.1998.0832
|
| [137] | Bhatnagar BS, Bogner RH, Pikal MJ (2007) Protein stability during freezing: separation of stresses and mechanisms of protein stabilization. Pharm Dev Technol 12: 505-523. |
| [138] | Jain NK, Roy I (2009) Effect of trehalose on protein structure. Protein Sci 18: 24-36. |
| [139] |
Capelle MA, Gurny R, Arvinte T (2007) High throughput screening of protein formulation stability: practical considerations. Eur J Pharm Biopharm 65: 131-148. doi: 10.1016/j.ejpb.2006.09.009
|
| [140] |
Kamerzell TJ, Esfandiary R, Joshi SB, et al. (2011) Protein-excipient interactions: mechanisms and biophysical characterization applied to protein formulation development. Adv Drug Deliv Rev 63: 1118-1159. doi: 10.1016/j.addr.2011.07.006
|
| [141] |
Ohtake S, Kita Y, Arakawa T (2011) Interactions of formulation excipients with proteins in solution and in the dried state. Adv Drug Deliv Rev 63: 1053-1073. doi: 10.1016/j.addr.2011.06.011
|
| [142] | Porfire AS, Tomuţa I, Irache JM, et al. (2009) The influence of the formulation factors on physico-chemical properties of dextran associated Gantrez an nanoparticles. Farmacia 57: 463-472. |
| [143] | Torres-Lugo M, Peppas NA (1999) Molecular design and in vitro studies of novel pH-sensitive hydrogels for the oral delivery of calcitonin. Macromolecules 32: 6646-6651. |
| [144] |
Szelke H, Schübel S, Harenberg J, et al. (2010) Interaction of heparin with cationic molecular probes: probe charge is a major determinant of binding stoichiometry and affinity. Bioorg Med Chem Lett 20: 1445-1447. doi: 10.1016/j.bmcl.2009.12.105
|
| [145] |
Houska M, Brynda E (1997) Interactions of proteins with polyelectrolytes at solid/liquid interfaces: Sequential adsorption of albumin and heparin. J Colloid Interface Sci 188: 243-250. doi: 10.1006/jcis.1996.4576
|
| [146] |
Guo B, Anzai J, Osa T (1996) Adsorption behavior of serum albumin on electrode surfaces and the effects of electrode potential. Chem Pharm Bull (Tokyo) 44: 800-803. doi: 10.1248/cpb.44.800
|
| [147] |
Langer K, Balthasar S, Vogel V, et al. (2003) Optimization of the preparation process for human serum albumin (HSA) nanoparticles. Int J Pharm 257: 169-180. doi: 10.1016/S0378-5173(03)00134-0
|
| [148] |
Prestrelski SJ, Pikal KA, Arakawa T (1995) Optimization of lyophilization conditions for recombinant human interleukin-2 by dried-state conformational analysis using fourier-transform infrared spectroscopy. Pharm Res 12: 1250-1259. doi: 10.1023/A:1016296801447
|
| [149] |
van Dijkhuizen-Radersma R, Nicolas HM, van de Weert M, et al. (2002) Stability aspects of salmon calcitonin entrapped in poly(ether-ester) sustained release systems. Int J Pharm 248: 229-237. doi: 10.1016/S0378-5173(02)00458-1
|
| [150] |
Baudys M, Mix D, Kim SW (1996) Stabilization and intestinal absorption of human calcitonin. J Control Rel 39: 145-151. doi: 10.1016/0168-3659(95)00148-4
|
| [151] |
Sigurjónsdóttir JF, Loftsson T, Másson M (1999) Influence of cyclodextrins on the stability of the peptide salmon calcitonin in aqueous solution. Int J Pharm 186: 205-213. doi: 10.1016/S0378-5173(99)00183-0
|
| [152] |
Mueller C, Capelle MA, Arvinte T, et al. (2011) Noncovalent pegylation by dansyl-poly(ethylene glycol)s as a new means against aggregation of salmon calcitonin. J Pharm Sci 100: 1648-1662. doi: 10.1002/jps.22401
|
| [153] |
Mueller C, Capelle MA, Arvinte T, et al. (2011) Tryptophan-mPEGs: novel excipients that stabilize salmon calcitonin against aggregation by non-covalent PEGylation. Eur J Pharm Biopharm 79: 646-657. doi: 10.1016/j.ejpb.2011.06.003
|
| [154] |
Mueller C, Capelle MA, Seyrek E, et al. (2012) Noncovalent PEGylation: different effects of dansyl-, L-tryptophan-, phenylbutylamino-, benzyl- and cholesteryl-PEGs on the aggregation of salmon calcitonin and lysozyme. J Pharm Sci 101: 1995-2008. doi: 10.1002/jps.23110
|
| [155] |
Remmele RL, Krishnan S, Callahan WJ (2012) Development of stable lyophilized protein drug products. Curr Pharm Biotechnol 13: 471-496. doi: 10.2174/138920112799361990
|
| [156] |
Kasper JC, Friess W (2011) The freezing step in lyophilization: physico-chemical fundamentals, freezing methods and consequences on process performance and quality attributes of biopharmaceuticals. Eur J Pharm Biopharm 78: 248-263. doi: 10.1016/j.ejpb.2011.03.010
|
| [157] | Costantino HR, Pikal MJ (2004) Lyophilization of Biopharmaceuticals. Virginia: AAPS Press. |
| [158] |
Susi H, Byler DM (1983) Protein structure by Fourier transform infrared spectroscopy: second derivative spectra. Biochem Biophys Res Commun 115: 391-397. doi: 10.1016/0006-291X(83)91016-1
|
| [159] |
Prestrelski SJ, Tedeschi N, Arakawa T, et al. (1993) Dehydration-induced conformational transitions in proteins and their inhibition by stabilizers. Biophys J 65: 661-671. doi: 10.1016/S0006-3495(93)81120-2
|
| [160] |
Lee HE, Lee MJ, Park CR, et al. (2010) Preparation and characterization of salmon calcitonin-sodium triphosphate ionic complex for oral delivery. J Control Rel 143: 251-257. doi: 10.1016/j.jconrel.2009.12.011
|
| [161] |
Steckel H, Brandes HG (2004) A novel spray-drying technique to produce low density particles for pulmonary delivery. Int J Pharm 278: 187-195. doi: 10.1016/j.ijpharm.2004.03.010
|
| [162] |
Seville PC, Li HY, Learoyd TP (2007) Spray-dried powders for pulmonary drug delivery. Crit Rev Ther Drug Carrier Syst 24: 307-360. doi: 10.1615/CritRevTherDrugCarrierSyst.v24.i4.10
|
| [163] | Sollohub K, Cal K (2010) Spray drying technique: II. Current applications in pharmaceutical technology. J Pharm Sci 99: 587-597. |
| [164] | Telko M, Hickey A (2005) Dry powder inhaler formulation. Respir Care 50: 1209-1227. |
| [165] |
Ameri M, Maa Y (2006) Spray Drying of Biopharmaceuticals: Stability and Process Considerations. Drying Technol: A Int J 24: 763-768. doi: 10.1080/03602550600685275
|
| [166] |
Chan HK, Clark AR, Feeley JC, et al. (2004) Physical stability of salmon calcitonin spray-dried powders for inhalation. J Pharm Sci 93: 792-804. doi: 10.1002/jps.10594
|
| [167] | Yang M, Velaga S, Yamamoto H, et al. (2007) Characterisation of salmon calcitonin in spray-dried powder for inhalation. Effect of chitosan. Int J Pharm 331: 176-181. |
| [168] | Sinsuebpol C, Chatchawalsaisin J, Kulvanich P (2013) Preparation and in vivo absorption evaluation of spray dried powders containing salmon calcitonin loaded chitosan nanoparticles for pulmonary delivery. Drug Des Devel Ther 7: 861-873. |
| [169] | Amaro MI, Tewes F, Gobbo O, et al. (2015) Formulation, stability and pharmacokinetics of sugar-based salmon calcitonin-loaded nanoporous/nanoparticulate microparticles (NPMPs) for inhalation. Int J Pharm 483: 6-18. |
| [170] |
Lechuga-Ballesteros D, Charan C, Stults CL, et al. (2008) Trileucine improves aerosol performance and stability of spray-dried powders for inhalation. J Pharm Sci 97: 287-302. doi: 10.1002/jps.21078
|
| [171] |
Tewes F, Gobbo OL, Amaro MI, et al. (2011) Evaluation of HP βCD-PEG microparticles for salmon calcitonin administration via pulmonary delivery. Mol Pharm 8: 1887-1898. doi: 10.1021/mp200231c
|
| [172] | Epand RM, Epand RF, Orlowski RC, et al. (1983) Amphipathic helix and its relationship to the interaction of calcitonin with phospholipids. Biochemistry 22: 5074-5084. |
| [173] | Epand RM, Epand RF (1986) Conformational flexibility and biological activity of salmon calcitonin. Biochemistry 25: 1964-1968. |
| [174] |
Green FR 3rd, Lynch B, Kaiser ET (1987) Biological and physical properties of a model calcitonin containing a glutamate residue interrupting the hydrophobic face of the idealized amphiphilic alpha-helical region. Proc Natl Acad Sci U S A 84: 8340-8344. doi: 10.1073/pnas.84.23.8340
|
| [175] |
Nabuchi Y, Asoh Y, Takayama M (2004) Folding analysis of hormonal polypeptide calcitonins and the oxidized calcitonins using electrospray ionization mass spectrometry combined with H/D exchange. J Am Soc Mass Spectrom 15: 1556-1564. doi: 10.1016/j.jasms.2004.07.007
|
| [176] |
Siligardi G, Samorí B, Melandri S, et al. (1994) Correlations between biological activities and conformational properties for human, salmon, eel, porcine calcitonins and Elcatonin elucidated by CD spectroscopy. Eur J Biochem 221: 1117-1125. doi: 10.1111/j.1432-1033.1994.tb18832.x
|
| [177] |
Andreotti G, Méndez BL, Amodeo P, et al. (2006) Structural determinants of salmon calcitonin bioactivity: the role of the Leu-based amphipathic alpha-helix. J Biol Chem 281: 24193-24203. doi: 10.1074/jbc.M603528200
|
| [178] |
Stefani M (2004) Protein misfolding and aggregation: new examples in medicine and biology of the dark side of the protein world. Biochim Biophys Acta 1739: 5-25. doi: 10.1016/j.bbadis.2004.08.004
|
| [179] |
Renukuntla J, Vadlapudi AD, Patel A, et al. (2013) Approaches for enhancing oral bioavailability of peptides and proteins. Int J Pharm 447: 75-93. doi: 10.1016/j.ijpharm.2013.02.030
|
| [180] |
Choonara BF, Choonara YE, Kumar P, et al. (2014) A review of advanced oral drug delivery technologies facilitating the protection and absorption of protein and peptide molecules. Biotechnol Adv 32: 1269-1282. doi: 10.1016/j.biotechadv.2014.07.006
|
| [181] |
Smart AL, Gaisford S, Basit AW (2014) Oral peptide and protein delivery: intestinal obstacles and commercial prospects. Expert Opin Drug Deliv 11: 1323-1335. doi: 10.1517/17425247.2014.917077
|
| [182] | Franks F, Hatley RHM, Mathias SF (1991) Materials science and the production of shelf-stable biologicals. Pharm Technol Int 3: 24-34. |
| [183] |
Slade L, Levine H (1991) Beyond water activity: Recent advances based on an alternative approach to the assessment of food quality and safety. Crit Rev Food Sci Nutri 30: 115-360. doi: 10.1080/10408399109527543
|
| [184] | Carpenter JF, Prestrelski SJ, Arakawa T (1993) Separation of freezing- and drying-induced denaturation of lyophilized proteins using stress-specific stabilization. I. Enzyme activity and calorimetric studies. Arch Biochem Biophys 303: 456-464. |
| [185] |
Carpenter JF, Crowe JH (1989) An infrared spectroscopic study of the interactions of carbohydrates with dried proteins. Biochemistry 28: 3916-3922. doi: 10.1021/bi00435a044
|
| [186] | Crowe JH, Crowe LM, Carpenter JF (1993) Preserving dry biomaterials: The water replacement hypothesis, Part 1. BioPharm 6: 28-29, 32-33. |
| [187] | Crowe JH, Crowe LM, Carpenter JF (1993) Preserving dry biomaterials: the water replacement hypothesis, Part 2. BioPharm 6: 40-43. |
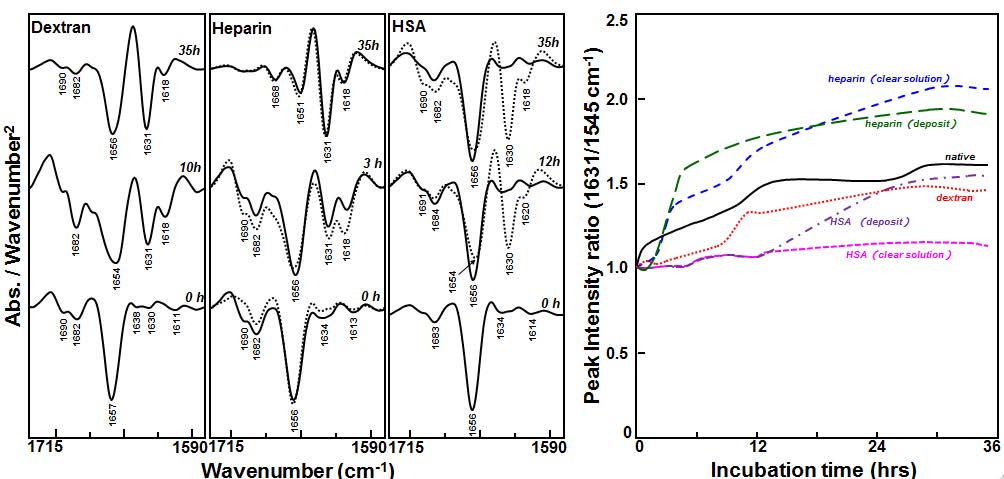
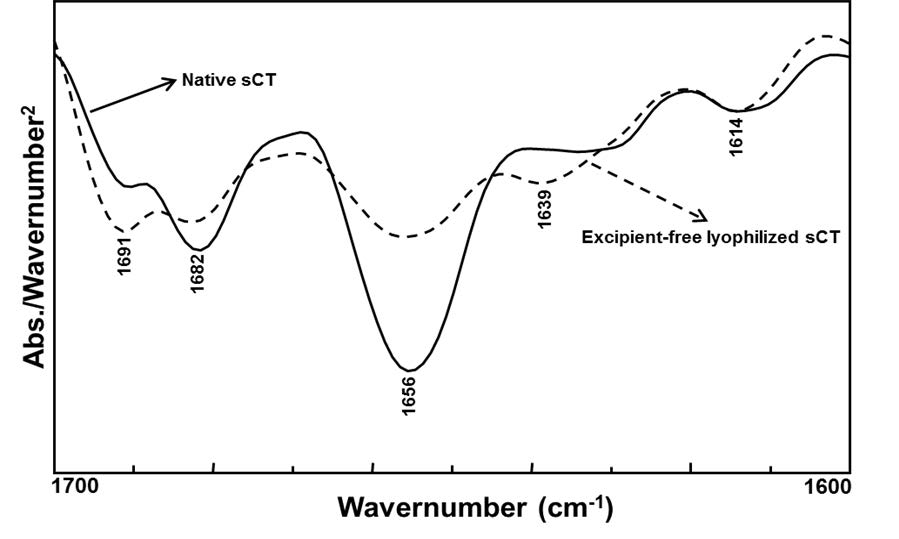
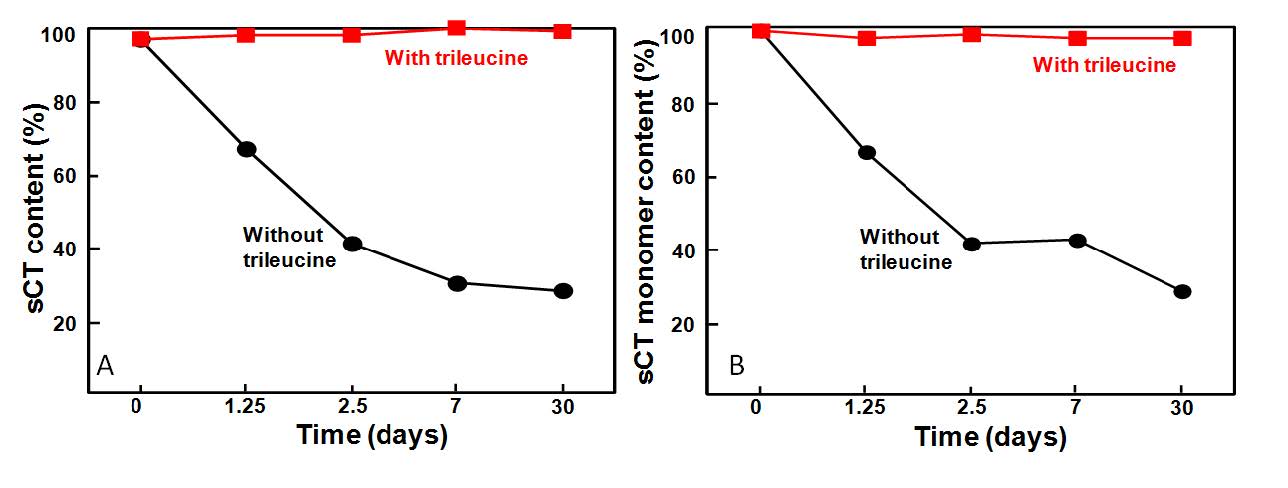
Figures(11) / Tables(2)

Shan-Yang Lin. Salmon calcitonin: conformational changes and stabilizer effects[J]. AIMS Biophysics, 2015, 2(4): 695-723. doi: 10.3934/biophy.2015.4.695







 DownLoad:
DownLoad: