Citation: Hayet Houmani, Francisco J Corpas. Differential responses to salt-induced oxidative stress in three phylogenetically related plant species: Arabidopsis thaliana (glycophyte), Thellungiella salsuginea and Cakile maritima (halophytes). Involvement of ROS and NO in the control of K+/Na+ homeostasis[J]. AIMS Biophysics, 2016, 3(3): 380-397. doi: 10.3934/biophy.2016.3.380

| [1] |
Kader MA, Lindberg S (2010) Cytosolic calcium and pH signaling in plants under salinity stress. Plant Signal Behav 5: 233–238. doi: 10.4161/psb.5.3.10740

|
| [2] |
Wang Y, Wu WH (2013) Potassium transport and signaling in higher plants. Annu Rev Plant Biol 64: 451–76. doi: 10.1146/annurev-arplant-050312-120153
|
| [3] |
Munns R, James RA, Läuchli A (2006) Approaches to increasing the salt tolerance of wheat and other cereals. J Exp Bot 57: 1025–1043. doi: 10.1093/jxb/erj100
|
| [4] |
Kader MA (2010) Cytosolic calcium and pH signaling in plants under salinity stress. Plant Signal Behav. 5: 233–238. doi: 10.4161/psb.5.3.10740
|
| [5] |
Hasegawa PM, Bressan RA, Zhu JK, et al. (2000). Plant cellular and molecular responses to high salinity. Annu Rev Plant Physiol Plant Mol Biol 51: 463–499. doi: 10.1146/annurev.arplant.51.1.463
|
| [6] |
Tester M, Davenport R (2003) Na+ tolerance and Na+ transport in higher plants. Ann Bot 91: 503–527. doi: 10.1093/aob/mcg058
|
| [7] | Newell N (2013) Review: Effects of Soil Salinity on Plant Growth. Plant Physiol. |
| [8] | Shabala S, Pottosin II (2010) Potassium and potassium-permeable channels in plant salt tolerance. Signal Commun Plants 2010: 87–110. |
| [9] |
Jiang C, Belfield EJ, Cao Y, et al. (2013) An Arabidopsis Soil-Salinity-Tolerance Mutation Confers Ethylene-Mediated Enhancement of Sodium/Potassium homeostasis. Plant Cell 25: 3535–3552. doi: 10.1105/tpc.113.115659
|
| [10] |
Zhu JK (2003) Regulation of ion homeostasis under salt stress. Curr Opin Plant Biol 6: 441–445. doi: 10.1016/S1369-5266(03)00085-2
|
| [11] |
Rubio F, Gassmann W, Schroeder JI (1995) Sodium-driven potassium uptake by the plant potassium transporter HKT1 and mutations conferring salt tolerance. Science 270: 1660–1663. doi: 10.1126/science.270.5242.1660
|
| [12] |
Uozumi N, Kim EJ, Rubio F, et al. (2000) The Arabidopsis HKT1 Gene Homolog Mediates Inward Na+ Currents in Xenopus laevis Oocytes and Na+ Uptake in Saccharomyces cerevisiae. Plant Physiol 122: 1249–1259. doi: 10.1104/pp.122.4.1249
|
| [13] |
Venema K, Quintero FJ, Pardo JM, et al. (2002) The Arabidopsis Na+/H+ exchanger AtNHX1 catalyzes low affinity Na+ and K+ transport in reconstituted liposomes. J Biol Chem 277: 2413–2418. doi: 10.1074/jbc.M105043200
|
| [14] |
Maathuis FJM (2009) Physiological functions of mineral macronutrients. Curr Opin Plant Biol 12: 250–258. doi: 10.1016/j.pbi.2009.04.003
|
| [15] |
Santa-Maria GE, Rubio F, Dubcovsky J, et al. (1997) The HAK1 gene of barley is a member of a large gene family and encodes a high-affinity potassium transporter. The Plant Cell 9: 2281–2289. doi: 10.1105/tpc.9.12.2281
|
| [16] | Fu HH, Luan S (1998) AtKuP1: a dual-affinity K+ transporter from Arabidopsis. Plant Cell 10: 63–73. |
| [17] |
Nieves-Cordones M, Aleman F, MartinezV, et al. (2010) The Arabidopsis thaliana HAK5 K+ transporter is required for plant growth and K+ acquisition from low K+ solutions under saline conditions. Mol Plant 3: 326–333. doi: 10.1093/mp/ssp102
|
| [18] |
Rubio F, Santa-Maria GE, Rodríguez-Navarro A (2000) Cloning of Arabidopsis and barley cDNAs encoding HAK potassium transporters in root and shoot cells. Physiol Plant 109: 34–43. doi: 10.1034/j.1399-3054.2000.100106.x
|
| [19] |
Qi Z, Hampton CR, Shin R, et al. (2008) The high affinity K+ transporter AtHAK5 plays a physiological role in planta at very low K+ concentrations and provides a caesium uptake pathway in Arabidopsis. J Exp Bot 59: 595–607. doi: 10.1093/jxb/erm330
|
| [20] |
Gorham J, Bridgers J. Dubcovsky J, et al. (1997) Genetic analysis and physiology of a trait for enhanced K+/Na+ discrimination in Wheat. New Phytol 137: 109–116. doi: 10.1046/j.1469-8137.1997.00825.x
|
| [21] |
Horie T, Sugawara M, Okada T, et al. (2011) Rice sodium-insensitive potassium transporter, OsHAK5, confers increased salt tolerance in tobacco BY2 cells. J Biosci Bioeng 111: 346–356. doi: 10.1016/j.jbiosc.2010.10.014
|
| [22] | Alemán F, Caballero F, Ródenas R et al. (2014) The F130S point mutation in the Arabidopsis high affinity K+ transporter AtHAK5 increases K+ over Na+ and Cs+ selectivity and confers Na+ and Cs+ tolerance to yeast under heterologous expression. Front Plant Sci 5: 430. |
| [23] |
Shabala S, CuinTA (2008) Potassium transport and plant salt tolerance. Physiol Plant 133: 651–669. doi: 10.1111/j.1399-3054.2007.01008.x
|
| [24] |
Haro R, Bañuelos MA, Rodriguez‐Navarro A (2010) High‐affinity sodium uptake in land plants. Plant Cell Physiol 51: 68–79. doi: 10.1093/pcp/pcp168
|
| [25] |
Kronzucker HJ, Britto DT (2011) Sodium transport in plants: A critical review. New Phytol 189: 54–81. doi: 10.1111/j.1469-8137.2010.03540.x
|
| [26] |
Kronzucker HJ, Coskun D, Schulze LM et al. (2013) Sodium as nutrient and toxicant. Plant Soil 369: 1–23. doi: 10.1007/s11104-013-1801-2
|
| [27] |
Adams E, Shin R (2014) Transport, signaling, and homeostasis of potassium and sodium in plants. J Integr Plant Biol 56: 231–249. doi: 10.1111/jipb.12159
|
| [28] |
Demidchik V, Maathuis FJM (2007) Physiological roles of nonselective cation channels in plants: from salt stress to signalling and development. New Phytol 175: 387–404. doi: 10.1111/j.1469-8137.2007.02128.x
|
| [29] |
Sun J, Dai S, Wang R, et al. (2009) Calcium mediates root K+/Na+ homeostasis in poplar species differing in salt tolerance. Tree Physiol 29: 1175–1186. doi: 10.1093/treephys/tpp048
|
| [30] |
Gattward JN, Almeida AA, Souza JO, et al. (2012) Sodium‐potassium synergism in Theobroma cacao: Stimulation of photosynthesis, water‐use efficiency and mineral nutrition. Physiol Plant 146: 350–362. doi: 10.1111/j.1399-3054.2012.01621.x
|
| [31] | Schulze LM, Britto DT, Li M, et al. (2012) A pharmacological analysis of high-affinity sodium transport in barley (Hordeum vulgare L.): a 24Na+/42K+ study. J Exp Bot 63: 2479–2489. |
| [32] |
Zhu JK (2002) Salt and drought stress signal transduction in plants. Annu Rev Plant Biol 53: 247–273. doi: 10.1146/annurev.arplant.53.091401.143329
|
| [33] |
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Annu Rev Plant Biol 59: 651–681. doi: 10.1146/annurev.arplant.59.032607.092911
|
| [34] | Jayakannan M, Bose J, Babourina O, et al. (2013) Salicylic acid improves salinity tolerance in Arabidopsis by restoring membrane potential and preventing salt-induced K+ loss via a GORK channel. J Exp Bot 64: 2255–2268. |
| [35] |
Römheld V, Kirkby E (2010) Research on potassium in agriculture: needs and prospects. Plant Soil 335: 155–180. doi: 10.1007/s11104-010-0520-1
|
| [36] |
Debez A, Ben Rejeb K, Ghars MA, et al. (2013) Ecophysiological and genomic analysis of salt tolerance of Cakile maritima. Environ Exp Bot 92: 64–72. doi: 10.1016/j.envexpbot.2012.12.002
|
| [37] |
Zhu JK, Liu J, Xiong L (1998) Genetic analysis of salt tolerance in Arabidopsis: Evidence for a critical role to potassium nutrition. Plant Cell 10: 1181–1191. doi: 10.1105/tpc.10.7.1181
|
| [38] | Keutgen AJ, Pawelzik E (2008) Impacts of NaCl stress on plant growth and mineral nutrient assimilation in two cultivars of strawberry. Environ Exp Bot 65: 170–176. |
| [39] | Bassil E, Coku A, Blumwald E (2012) Cellular ion homeostasis: emerging roles of intracellular NHX Na+/H+ antiporters in plant growth and development. J Exp Bot 63: 5727–5740. |
| [40] |
Maathuis FJM, Amtmann A (1999) K+ nutrition and Na+ toxicity: The basis of cellular K+/Na+ ratios. Ann Bot 84: 123–133. doi: 10.1006/anbo.1999.0912
|
| [41] |
Flowers TJ, Galal HK, Bromham L (2010) Evolution of halophytes: multiple origins of salt tolerance in land plants. Funct Plant Biol 37: 604–612. doi: 10.1071/FP09269
|
| [42] |
Ali Z, Park HC, Ali A, et al. (2012) TsHKT1;2, a HKT1 Homolog from the Extremophile Arabidopsis Relative Thellungiella salsuginea, Shows K+ Specificity in the Presence of NaCl. Plant Physiol 158: 1463–1474. doi: 10.1104/pp.111.193110
|
| [43] | Wang M, Zheng Q, Shen Q, et al. (2013) The Critical Role of Potassium in Plant Stress Response. Int J Mol Sci14: 7370–7390. |
| [44] | Smith A, Jain A, Deal R, et al. (2010). Histone H2A.Z regulates the expression of several classes of phosphate starvation response genes, but not as a transcriptional activator. Plant Physiol 152: 217–225. |
| [45] |
Ardie SW, Liu S, Takano T (2010) Expression of the AKT1-type K+ channel gene from Puccinellia tenuiflora, PutAKT1, enhances salt tolerance in Arabidopsis. Plant Cell Rep 29: 865–874. doi: 10.1007/s00299-010-0872-2
|
| [46] |
Coskun D, Britto DT, Li MY, et al. (2013) Capacity and plasticity of potassium channels and high-affinity transporters in roots of barley and Arabidopsis. Plant Physiol 162: 496–511. doi: 10.1104/pp.113.215913
|
| [47] |
Sussman MR (1994) Molecular analysis of protein in the plant plasma membrane. Annu Rev Plant Physiol Plant Mol Biol 45: 211–234. doi: 10.1146/annurev.pp.45.060194.001235
|
| [48] | Blumwald E, Aharon GS, Apse MP (2000) Sodium transport in plant cells. Biochim Biophys Acta 1465: 140–151. |
| [49] | Gaxiola RA, Rao R, Sherman A, et al. (1999) The Arabidopsis thaliana proton transporters AtNhx1 and Avp1 can function in cation detoxification in yeast. Proc Natl Acad Sci USA 96: 1480–1485. |
| [50] |
Deinlein U, Stephan AB, Horie T, et al. (2014). Plant salt-tolerance mechanisms. Trends Plant Sci 19: 371–379. doi: 10.1016/j.tplants.2014.02.001
|
| [51] |
Tapken D, Hollmann M (2008) Arabidopsis thaliana glutamate receptor ion channel function demonstrated by ion pore transplantation. J Mol Biol 383: 36–48. doi: 10.1016/j.jmb.2008.06.076
|
| [52] | Shabala S, Wu H, Bose J (2015) Salt stress sensing and early signalling events in plant roots: Current knowledge and hypothesis. Plant Sci 241: 109–119. |
| [53] | Wegner, LH, De Boer AH (1997) Properties of two outward-rectifying channels in root xylem parenchyma cells suggest a role in K+ homeostasis and long-distance signaling. Plant Physiol 115: 1707–1719. |
| [54] |
Gaymard F, Pilot G, Lacombe B, et al. (1998) Identification and disruption of a plant shaker-like outward channel involved in K+ release into the xylem sap. Cell 94: 647–655. doi: 10.1016/S0092-8674(00)81606-2
|
| [55] |
Ren, Z.H, Gao JP, Li LG, et al. (2005) A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat Genet 37: 1141–1146. doi: 10.1038/ng1643
|
| [56] |
Yang T, Zhang S, Hu Y, et al. (2014) The role of a potassium transporter OsHAK5 in potassium acquisition and transport from roots to shoots in rice at low potassium supply levels. Plant Physiol 166: 945–959. doi: 10.1104/pp.114.246520
|
| [57] |
Benito B, Garciadeblas B, Rodriguez-Navarro A (2012) HAK transporters from Physcomitrella patens and Yarrowia lipolytica mediate sodium uptake. Plant Cell Physiol 53: 1117–1123. doi: 10.1093/pcp/pcs056
|
| [58] |
WU SJ, Ding L, Zhu JK (1996) SOS1, a genetic locus essential for salt tolerance and potassium acquisition. Plant Cell 8: 617–627. doi: 10.1105/tpc.8.4.617
|
| [59] |
Oh DH, Gong QQ, Ulanov A, et al. (2007) Sodium stress in the halophyte Thellungiella halophila and transcriptional changes in a thsos1-RNA interference line. J Integr Plant Biol 49: 1484–1496. doi: 10.1111/j.1672-9072.2007.00548.x
|
| [60] |
Ahn SJ, Shin R, Schachtman DP (2004) Expression of KT/KUP genes in Arabidopsis and the role of root hairs in K+ Uptake. Plant Physiol 134: 1135–1145. doi: 10.1104/pp.103.034660
|
| [61] |
Berthomieu P, Conéjéro G, Nublat A, et al. (2003) Functional analysis of AtHKT1 in Arabidopsis shows that Na+ recirculation by the phloem is crucial for salt tolerance. EMBO J 22: 2004–2014. doi: 10.1093/emboj/cdg207
|
| [62] | Ali A, Raddatz N, Aman R, et al. (2016) A Single Amino Acid Substitution in the Sodium Transporter HKT1 Associated with Plant Salt Tolerance. Plant Physiol. 171: 2112–2126. |
| [63] |
Corratgé-Faillie C, Jabnoune M, Zimmermann S, et al. (2010) Potassium and sodium transport in non-animal cells: the Trk/Ktr/HKT transporter family. Cell Mol Life Sci 67: 2511–2532. doi: 10.1007/s00018-010-0317-7
|
| [64] |
Alemán F, Nieves-Cordones M, Martínez V, et al. (2011) Root K+ acquisition in plants: the Arabidopsis thaliana model. Plant Cell Physiol 52: 1603–1612. doi: 10.1093/pcp/pcr096
|
| [65] | Nieves-Cordones M, Alemán F, Martínez V, et al. (2014) K+ uptake in plant roots. the systems involved, their regulation and parallels in other organisms. J Plant Physiol 171: 688–695. |
| [66] |
Rubio F, Nieves-Cordones M, Alemán F, et al. (2008) Relative contribution of AtHAK5 and AtAKT1 to K+ uptake in the high-affinity range of concentrations. Physiol Plant 134: 598–608. doi: 10.1111/j.1399-3054.2008.01168.x
|
| [67] |
Rubio F, Alemán F, Nieves-Cordones M, et al. (2010) Studies on Arabidopsis athak5, atakt1 double mutants disclose the range of concentrations at which AtHAK5, AtAKT1 and unknown systems mediate K+ uptake. Physiol Plant 139: 220–228. doi: 10.1111/j.1399-3054.2010.01354.x
|
| [68] |
PyoYJ, Gierth M, Schroeder JI, et al. (2010) High-Affinity K+ transport in Arabidopsis: AtHAK5 and AKT1 are vital for seedling establishment and postgermination growth under low-potassium conditions. Plant Physiol 153: 863–875. doi: 10.1104/pp.110.154369
|
| [69] |
Alemán F, Nieves-Cordones M, Martínez V, et al. (2009). Differential regulation of the HAK5 genes encoding the high-affinity K+ transporters of Thellungiella halophila and Arabidopsis thaliana. Environ Exp Bot 65: 263–269. doi: 10.1016/j.envexpbot.2008.09.011
|
| [70] |
Adams E, Abdollahi P, Shin R (2013) Cesium inhibits plant growth through Jasmonate signaling in Arabidopsis thaliana. Int J Mol Sci 14: 4545–4559. doi: 10.3390/ijms14034545
|
| [71] |
Sun Y, Kong X, Li C et al. (2015) Potassium retention under salt stress is associated with natural variation in salinity tolerance among Arabidopsis accessions. PLoS One 10: e0124032. doi: 10.1371/journal.pone.0124032
|
| [72] |
Rodríguez-Rosales MP, Jiang XJ, Gálvez FJ, et al. (2008) Overexpression of the tomato K+/H+ antiporter LeNHX2 confers salt tolerance by improving potassium compartmentalization. New Phytol 179: 366–77. doi: 10.1111/j.1469-8137.2008.02461.x
|
| [73] | Leidi EO, Barragán V, Rubio L, et al. (2010) The AtNHX1 exchanger mediates potassium compartmentation in vacuoles of transgenic tomato. Plant J 61: 495–506. |
| [74] |
Ghars MA, Parre E, Debez A, et al. (2008) Comparative salt tolerance analysis between Arabidopsis thaliana and Thellungiella halophila, with special emphasis on K+/Na+ selectivity and proline accumulation. J Plant Physiol 165: 588–599. doi: 10.1016/j.jplph.2007.05.014
|
| [75] |
Bailey CD, Koch MA, Mayer M, et al. (2006) Toward a global phylogeny of the Brassicaceae. Mol Biol Evol 23: 2142–2160. doi: 10.1093/molbev/msl087
|
| [76] |
Debez A, Ben HK, Grigno C, et al. (2004) Salinity effects on germination, growth, and seed production of the halophyte Cakile maritime. Plant Soil 262: 179–189. doi: 10.1023/B:PLSO.0000037034.47247.67
|
| [77] |
Houmani H, Rodríguez-Ruiz M, Palma JM, et al. (2016). Modulation of superoxide dismutase (SOD) isozymes by organ development and high long-term salinity in the halophyte Cakile maritima. Protoplasma 253: 885–894. doi: 10.1007/s00709-015-0850-1
|
| [78] |
Debez A, Koyro HW, Grignon C, et al. (2008) Relationship between the photosynthetic activity and the performance of Cakile maritima after long-term salt treatment. Physiol Plantarum 133: 373–385. doi: 10.1111/j.1399-3054.2008.01086.x
|
| [79] | Demidchik V, Tester M (2002) Sodium Fluxes through Nonselective Cation Channels in the Plasma Membrane of Protoplasts from Arabidopsis Roots. Plant Physiol 128: 379–387. |
| [80] | Yadav S, Irfan M, Ahmad A, et al. (2011) Causes of salinity and plant manifestations to salt stress: A review. J Environ Biol 32: 667–685. |
| [81] |
Ellouzi H, Ben Hamed K, Hernández I, et al. (2014) A comparative study of the early osmotic, ionic, redox and hormonal signaling response in leaves and roots of two halophytes and a glycophyte to salinity. Planta 240: 1299–1317. doi: 10.1007/s00425-014-2154-7
|
| [82] |
Fan W, Zhang Z, Zhang Y (2009) Cloning and molecular characterization of fructose-1,6-bisphosphate aldolase gene regulated by high-salinity and drought in Sesuvium portulacastrum. Plant Cell Rep 28: 975–984. doi: 10.1007/s00299-009-0702-6
|
| [83] |
Debez A, Saadaoui D, Ramani B, et al. (2006). Leaf H+-ATPase activity and photosynthetic capacity of Cakile maritima under increasing salinity. Environ Exp Bot 57: 285–295. doi: 10.1016/j.envexpbot.2005.06.009
|
| [84] |
Volkov V, Wang B, Dominy PJ, et al. (2004) Thellungiella halophila, a salt-tolerant relative of Arabidopsis thaliana, possesses effective mechanisms to discriminate between potassium and sodium. Plant Cell Environ 27: 1–14. doi: 10.1046/j.0016-8025.2003.01116.x
|
| [85] |
Wang B, Davenport RJ, Volkov V, et al. (2006) Low unidirectional sodium influx into root cells restricts net sodium accumulation in Thellungiella halophila, a salt-tolerant relative of Arabidopsis thaliana. J Exp Bot 57: 1161–1170. doi: 10.1093/jxb/erj116
|
| [86] |
Wong CE, Li Y, Whitty BR, et al. (2005) Expressed sequence tags from the Yukon ecotype of Thellungiella reveal that gene expression in response to cold, drought and salinity shows little overlap. Plant Mol Biol 58: 561–574. doi: 10.1007/s11103-005-6163-6
|
| [87] |
Cuin TA, Betts SA, Chalmandrier R, et al. (2008). A root’s ability to retain K+ correlates with salt tolerance in wheat. J Exp Bot 59: 2697–2706. doi: 10.1093/jxb/ern128
|
| [88] |
Wang ZI, Li PH, Fredricksen M, et al. (2004) Expressed sequence tags from Thellungiella halophila, a new model to study plant salt-tolerance. Plant Sci 166: 609–616. doi: 10.1016/j.plantsci.2003.10.030
|
| [89] |
Vera-Estrella R, Barkla BJ, Garcia-Ramirez L, et al. (2005) Salt stress in Thellungiella halophila activates Na+ transport mechanisms required for salinity tolerance. Plant Physiol 139: 1507–1517. doi: 10.1104/pp.105.067850
|
| [90] |
Volkov V, Amtmann A (2006) Thellungiella halophila, a salt-tolerant relative of Arabidopsis thaliana, has specific root ion-channel features supporting K+ /Na+ homeostasis under salinity stress. Plant J 48: 342–353. doi: 10.1111/j.1365-313X.2006.02876.x
|
| [91] | Volkov V, Wang B, Dominy PJ, et al. (2003) Thellungiella halophila, a salt-tolerant relative of Arabidopsis thaliana, possesses effective mechanisms to discriminate between potassium and sodium. Plant Cell Environ 27: 1–14. |
| [92] | Kosová K, Prášil TI, Vítámvás P (2013) Protein contribution to plant salinity response and tolerance acquisition. Int J Mol Sci< 14: 6757–6789. |
| [93] |
Maathuis FJM (2014) Sodium in plants: perception, signaling, and regulation of sodium fluxes. J Exp Bot 65: 849–858. doi: 10.1093/jxb/ert326
|
| [94] | Ellouzi H, Ben Hamed K, Cela J, et al. (2011) Early effects of salt stress on the physiological and oxidative status of Cakile maritima (halophyte) and Arabidopsis thaliana (glycophyte). Physiol Plant 142: 128–43. |
| [95] |
Horie T, Hauser F, Schroeder JI (2009) HKT transporter-mediated salinity resistance mechanisms in Arabidopsis and monocot crop plants. Trends Plant Sci 14: 660–668. doi: 10.1016/j.tplants.2009.08.009
|
| [96] |
Rodríguez-Rosales MP, Gálvez FJ, Huertas R et al. (2009). Plant NHX cation/proton antiporters. Plant Signal Behav 4: 265–276. doi: 10.4161/psb.4.4.7919
|
| [97] |
Han M, Wu W, Wu WH, et al. (2016) Potassium Transporter KUP7 Is Involved in K(+) Acquisition and Translocation in Arabidopsis Root under K(+)-Limited Conditions. Mol Plant 9: 437–446. doi: 10.1016/j.molp.2016.01.012
|
| [98] | Trono D, Laus MN, Soccio M, et al. (2015) Modulation of potassium channel activity in the balance of ROS and ATP production by durum wheat mitochondria-an amazing defense tool against hyperosmotic stress. Front Plant Sci 6: 1072. |
| [99] |
Sun J, Wang MJ, Ding MQ et al. (2010) H2O2 and cytosolic Ca2+ signals triggered by the PM H+-coupled transport system mediate K+/Na+ homeostasis in NaCl-stressed Populus euphratica cells. Plant Cell Environ 33: 943–958. doi: 10.1111/j.1365-3040.2010.02118.x
|
| [100] |
Jung JY, Shin R, Schachtman DP (2009) Ethylene mediates response and tolerance to potassium deprivation in Arabidopsis. Plant Cell 21: 607–621. doi: 10.1105/tpc.108.063099
|
| [101] |
Shin R, Schachtman DP (2004) Hydrogen peroxide mediates plant root cell response to nutrient deprivation. Proc Natl Acad Sci USA 101: 8827–8832. doi: 10.1073/pnas.0401707101
|
| [102] |
Garcia-Mata C, Wang J ,Gajdanowicz P, et al. (2010) A Minimal Cysteine Motif Required to Activate the SKOR K Channel of Arabidopsis by the Reactive Oxygen Species H2O2. J Biol Chem 285: 29286–29294. doi: 10.1074/jbc.M110.141176
|
| [103] | Ho CH, YF tsay (2010) Nitrate, ammonium, and potassium sensing and signaling. Curr Opin< Plant Biol 13: 604–610. |
| [104] |
Kim MJ, Ciani S, Schachtman DP (2010) A peroxidase contributes to ROS production during Arabidopsis root response to potassium deficiency. Mol Plant 3: 420–427. doi: 10.1093/mp/ssp121
|
| [105] |
Alscher RG, Erturk N, Heath LS (2002) Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J Exp Bot 53: 1331–1341. doi: 10.1093/jexbot/53.372.1331
|
| [106] |
Tsugane K, Kobayashi K, Niwa Y, et al. (1999) A recessive Arabidopsis mutant that grows photoautotrophically under salt stress shows enhanced active oxygen detoxification. Plant Cell 11: 1195–1206. doi: 10.1105/tpc.11.7.1195
|
| [107] |
M’rah S, Ouerghi Z, Berthomieu C, et al. (2006) Effects of NaCl on the growth, ion accumulation and photosynthetic parameters of Thellungiella halophila. J Plant Physiol 163: 1022–1031. doi: 10.1016/j.jplph.2005.07.015
|
| [108] |
Ellouzi H, Ben Hamed K, Asensi-Fabado MA, et al. (2013) Drought and cadmium may be as effective as salinity in conferring subsequent salt stress tolerance in Cakile maritima. Planta 237: 1311–1323. doi: 10.1007/s00425-013-1847-7
|
| [109] |
Ma L, Zhang H, Sun L, et al. (2012) NADPH oxidase AtrbohD and AtrbohF function in ROS-dependent regulation of Na+/K+homeostasis in Arabidopsis under salt stress. J Exp Bot 63: 305–317. doi: 10.1093/jxb/err280
|
| [110] |
Bose J, Rodrigomoreno A, Shabala S (2014) ROS homeostasis in halophytes in thecontext of salinity stress tolerance. J Exp Bot 65: 1241–1257. doi: 10.1093/jxb/ert430
|
| [111] |
Corpas FJ, Barroso JB (2015) Nitric oxide from a "green" perspective. Nitric Oxide 45: 15–19. doi: 10.1016/j.niox.2015.01.007
|
| [112] |
Tanou G, Molassiotis A, Diamantidis G (2009) Hydrogen peroxide- and nitric oxide-induced systemic antioxidant prime-like activity under NaCl-stress and stress-free conditions in citrus plants. J Plant Physiol 166: 1904-1913. doi: 10.1016/j.jplph.2009.06.012
|
| [113] | Manai J, Kalai T, Gouia H, et al. (2014) Exogenous nitric oxide (NO) ameliorates salinity-induced oxidative stress in tomato (Solanum lycopersicum) plants. J Soil Sci Plant Nutr 14: 433–446. |
| [114] |
Crawford NM (2006) Mechanisms for nitric oxide synthesis in plants. J Exp Bot 57: 471–478. doi: 10.1093/jxb/erj050
|
| [115] | Gupta B, Huang B (2014) Mechanism of salinity tolerance in plants: physiological, biochemical, and molecular characterization. Int J Genomics 2014: 727–740. |
| [116] |
Xia J, Kong D, Xue S, et al. (2014) Nitric oxide negatively regulates AKT1-mediated potassium uptake through modulating vitamin B6 homeostasis in Arabidopsis. P Natl Acad Sci USA 111: 16196–16201. doi: 10.1073/pnas.1417473111
|
| [117] | Zhao MG, Tian QY, Zhang WH (2007) Nitric oxide synthase-dependent nitric oxide production is associated with salt tolerance in Arabidopsis. Plant Physiol< 144: 206–217. |
| [118] |
Chen J, Xiong DY, Wang WH, et al. (2013) Nitric oxide mediates root K+/Na+ balance in a mangrove plant, Kandelia obovata, by enhancing the expression of AKT1-type K+ channel and Na+/H+ antiporter under high salinity. PLoS One 8: 71543. doi: 10.1371/journal.pone.0071543
|
| [119] |
Kliebenstein DJ, Monde RA, Last RL (1998) Superoxide dismutase in Arabidopsis: an eclectic enzyme family with disparate regulation and protein localization. Plant Physiol. 118: 637–650. doi: 10.1104/pp.118.2.637
|
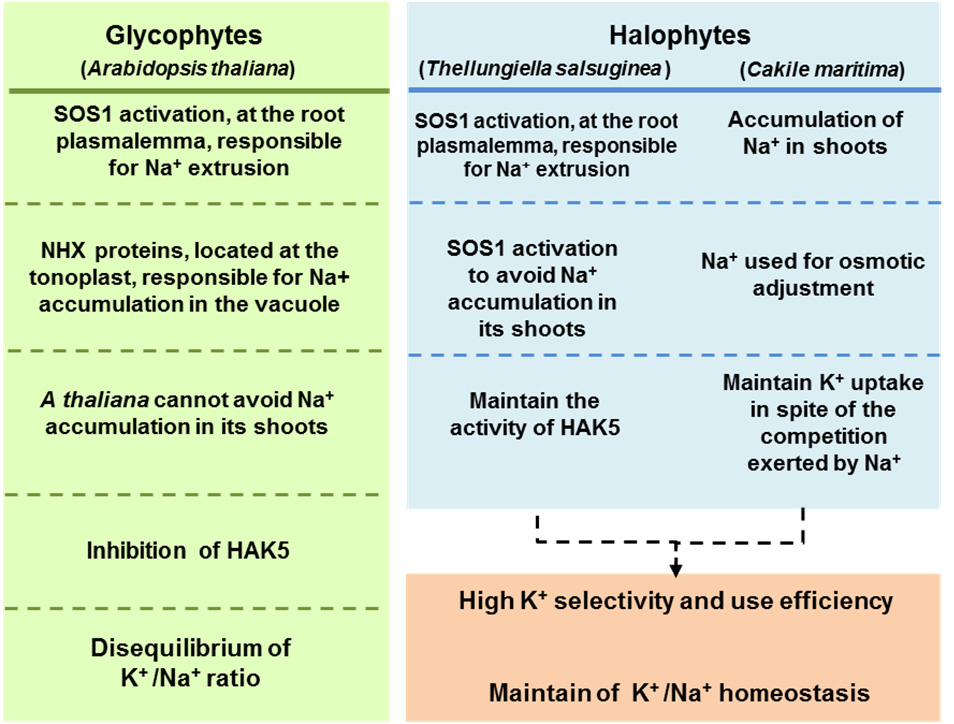
Figures(1) / Tables(2)

Hayet Houmani, Francisco J Corpas. Differential responses to salt-induced oxidative stress in three phylogenetically related plant species: Arabidopsis thaliana (glycophyte), Thellungiella salsuginea and Cakile maritima (halophytes). Involvement of ROS and NO in the control of K+/Na+ homeostasis[J]. AIMS Biophysics, 2016, 3(3): 380-397. doi: 10.3934/biophy.2016.3.380







 DownLoad:
DownLoad: